

автордың кітабын онлайн тегін оқу A Guide to the Study of Fishes, Volume 1 (of 2)
The cover image was created by the transcriber and is placed in the public domain.
GUIDE TO THE STUDY OF FISHES

[Pg ii] [Pg iii]
A GUIDE TO THE STUDY OF FISHES
BY
DAVID STARR JORDAN President of Leland Stanford Junior University
With Colored Frontispieces and 427 Illustrations
IN TWO VOLUMES Vol I.
"I am the wiser in respect to all knowledge and the better qualified for all fortunes for knowing that there is a minnow in that brook."—Thoreau

NEW YORK
HENRY HOLT AND COMPANY 1905
Copyright, 1905
BY
HENRY HOLT AND COMPANY
Published March, 1905
To
Theodore Gill,
Ichthyologist, Philosopher, Critic, Master in Taxonomy,
this volume is dedicated.
[Pg vi] [Pg vii]
PREFACE
This work treats of the fish from all the varied points of view of the different branches of the study of Ichthyology. In general all traits of the fish are discussed, those which the fish shares with other animals most briefly, those which relate to the evolution of the group and the divergence of its various classes and orders most fully. The extinct forms are restored to their place in the series and discussed along with those still extant.
In general, the writer has drawn on his own experience as an ichthyologist, and with this on all the literature of the science. Special obligations are recognized in the text. To Dr. Charles H. Gilbert, he is indebted for a critical reading of most of his proof-sheets; to Dr. Bashford Dean, for criticism of the proof-sheets of the chapters on the lower fishes; to Dr. William Emerson Ritter, for assistance in the chapters on Protochordata; to Dr. George Clinton Price, for revision of the chapters on lancelets and lampreys, and to Mr. George Clark, Secretary of Stanford University, for assistance of various kinds, notably in the preparation of the index. To Dr. Theodore Gill, he has been for many years constantly indebted for illuminating suggestions, and to Dr. Barton Warren Evermann, for a variety of favors. To Dr. Richard Rathbun, the writer owes the privilege of using illustrations from the "Fishes of North and Middle America" by Jordan and Evermann. The remaining plates were drawn for this work by Mary H. Wellman, Kako Morita, and Sekko Shimada. Many of the plates are original. Those copied from other authors are so indicated in the text.
No bibliography has been included in this work. A list of writers so complete as to have value to the student would make a volume of itself. The principal works and their authors are discussed in the chapter on the History of Ichthyology, and with this for the present the reader must be contented.
The writer has hoped to make a book valuable to technical students, interesting to anglers and nature lovers, and instructive to all who open its pages.
David Starr Jordan.
Palo Alto, Santa Clara County, Cal.,
October, 1904.
ERRATA[1]
VOL. I
- Frontispiece, for Paramia quinqueviltata read Paramia quinquevittata
- Page xiii, line 10, for Filefish read Tilefish
- 39, " 15, for Science read Sciences
- 52, lines 4 and 5, transpose hypocoracoid and hypercoracoid
- 115, line 24, for Hexagramidæ read Hexagrammidæ
- 162, " 7, The female salmon does as much as the male in covering the eggs.
- 169, last line, for immmediately read immediately
- 189, legend, for Miaki read Misaki
- 313, line 26, for sand-pits read sand-spits
- 322, " 7 and elsewhere, for Wood's Hole read Woods Hole
- 324, " 15, for Roceus read Roccus
- 327, " next to last, for masquinonqy read masquinongy
- 357, " 5, for Filefish read Tilefish
- 361, " 26, for 255 feet read 25 feet
- 368, " 26, for infallibility read fallibility
- 414, " 22, for West Indies read East Indies
- 419, " 23, for-99 read-96
- 420, " 28, for were read are
- 428, " 24, for Geffroy, St. Hilaire read Geoffroy St. Hilaire
- 428, " 25, for William Kitchener Parker read William Kitchen Parker
- 462, " 32, for Enterpneusta read Enteropneusta
[Pg x] [Pg xi]
CONTENTS
VOL. I.
CHAPTER I.
THE LIFE OF THE FISH (
Lepomis megalotis).
PAGE
What is a Fish?—The Long-eared Sunfish.—Form of the Fish.—Face of the Fish.—How the Fish Breathes.—Teeth of the Fish.—How the Fish Sees.—Color of the Fish.—The Lateral Line.—The Fins of the Fish.—The Skeleton of the Fish.—The Fish in Action.—The Air-bladder.—The Brain of the Fish.—The Fish's Nest.
3CHAPTER II.
THE EXTERIOR OF THE FISH.
Form of Body.—Measurement of the Fish.—The Scales or Exoskeleton.—Ctenoid and Cycloid Scales.—Placoid Scales.—Bony and Prickly Scales.—Lateral Line.—Function of the Lateral Line.—The Fins of Fishes.—Muscles.
16CHAPTER III.
THE DISSECTION OF THE FISH.
The Blue-green Sunfish.—The Viscera.—Organs of Nutrition.—The Alimentary Canal.—The Spiral Valve.—Length of the Intestine.
26CHAPTER IV.
THE SKELETON OF THE FISH.
Specialization of the Skeleton.—Homologies of Bones of Fishes.—Parts of the Skeleton.—Names of Bones of Fishes.—Bones of the Cranium.—Bones of the Jaws.—The Suspensorium of the Mandible.—Membrane Bones of Head.—Branchial Bones.—The Gill-arches.—The Pharyngeals.—The Vertebral Column.—The Interneurals and Interhæmals.—The Pectoral Limb.—The Shoulder-girdle.—The Posterior Limb.—Degeneration.—The Skeleton in Primitive Fishes.—The Skeleton of Sharks.—The Archipterygium.
34CHAPTER V.
MORPHOLOGY OF THE FINS OF FISHES.
Origin of the Fins of Fishes.—Origin of the Paired Fins.—Development of the Paired Fins in the Embryo.—Evidences of Palæontology.—Current Theories as to Origin of Paired Fin.—Balfour's Theory of the Lateral Fold.—Objections.—Objections to Gegenbaur's Theory.—Kerr's Theory of Modified External Gills.—Uncertain Conclusions.—Forms of the Tail in Fishes.—Homologies of the Pectoral Limb.—The Girdle in Fishes other than Dipnoans.
62CHAPTER VI.
THE ORGANS OF RESPIRATION.
How Fishes Breathe.—The Gill Structures.—The Air-bladder.—Origin of the Air-bladder.—The Origin of Lungs.—The Heart of the Fish.—The Flow of Blood.
91CHAPTER VII.
THE NERVOUS SYSTEM.
The Nervous System.—The Brain of the Fish.—The Pineal Organ.—The Brain of Primitive Fishes.—The Spinal Cord.—The Nerves.
109CHAPTER VIII.
THE ORGANS OF SENSE.
The Organs of Smell.—The Organs of Sight.—The Organs of Hearing.—Voices of Fishes.—The Sense of Taste.—The Sense of Touch.
115CHAPTER IX.
THE ORGANS OF REPRODUCTION.
The Germ-cells.—The Eggs of Fishes.—Protection of the Eggs.—Sexual Modification.
124CHAPTER X.
THE EMBRYOLOGY AND GROWTH OF FISHES.
Post-embryonic Development.—General Laws of Development.—The Significance of Facts of Development.—The Development of the Bony Fishes.—The Larval Development of Fishes.—Peculiar Larval Forms.—The Development of Flounders.—Hybridism.—The Age of Fishes.—Tenacity of
Life.—Effect of Temperature on Fishes.—Transportation of Fishes.—Reproduction of Lost Parts.—Monstrosities among Fishes.
131CHAPTER XI.
INSTINCTS, HABITS, AND ADAPTATIONS.
The Habits of Fishes.—Irritability of Animals.—Nerve-cells and Fibers.—The Brain or Sensorium.—Reflex Action.—Instinct.—Classification of Instincts.—Variability of Instincts.—Adaptations to Environment.—Flight of Fishes.—Quiescent Fishes.—Migratory Fishes.—Anadromous Fishes.—Pugnacity of Fishes.—Fear and Anger in Fishes.—Calling the Fishes.—Sounds of Fishes.—Lurking Fishes.—The Unsymmetrical Eyes of the Flounder.—Carrying Eggs in the Mouth.
152CHAPTER XII.
ADAPTATIONS OF FISHES.
Spines of the Catfishes.—Venomous Spines.—The Lancet of the Surgeon-fish.—Spines of the Sting-ray.—Protection through Poisonous Flesh of Fishes.—Electric Fishes.—Photophores or Luminous Organs.—Photophores in the Iniomous Fishes.—Photophores of Porichthys.—Globefishes.—Remoras.—Sucking-disks of Clingfishes.—Lampreys and Hogfishes.—The Swordfishes.—The Paddle-fishes.—The Sawfishes.—Peculiarities of Jaws and Teeth.—The Angler-fishes.—Relation of Number of Vertebræ to Temperature, and the Struggle for Existence.—Number of Vertebræ: Soft-rayed Fishes; Spiny-rayed Fishes; Fresh-water Fishes; Pelagic Fishes.—Variations in Fin-rays.—Relation of Numbers to Conditions of Life.—Degeneration of Structures.—Conditions of Evolution among Fishes.
179CHAPTER XIII.
COLORS OF FISHES.
Pigmentation.—Protective Coloration.—Protective Markings.—Sexual Coloration.—Nuptial Coloration.—Coral-reef Fishes.—Recognition Marks.—Intensity of Coloration.—Fading of Pigments in Spirits.—Variation in Pattern.
226CHAPTER XIV.
GEOGRAPHICAL DISTRIBUTION OF FISHES.
Zoogeography.—General Laws of Distribution.—Species Absent through Barriers.—Species Absent through Failure to Maintain Foothold.—Species Changed through Natural Selection.—Extinction of Species.—Barriers
Checking Movements of Marine Species.—Temperature the Central Fact in Distribution.—Agency of Ocean Currents.—Centers of Distribution.—Distribution of Marine Fishes.—Pelagic Fishes.—Bassalian Fishes.—Littoral Fishes.—Distribution of Littoral Fishes by Coast Lines.—Minor Faunal Areas.—Equatorial Fishes most Specialized.—Realms of Distribution of Fresh-water Fishes.—Northern Zone.—Equatorial Zone.—Southern Zone.—Origin of the New Zealand Fauna.
237CHAPTER XV.
ISTHMUS BARRIERS SEPARATING FISH FAUNAS.
The Isthmus of Suez.—The Fish Fauna of Japan.—Fresh-water Faunas of Japan.—Faunal Areas of Marine Fishes of Japan.—Resemblance of Japanese and Mediterranean Fish Faunas.—Significance of Resemblances.—Differences between Japanese and Mediterranean Fish Faunas.—Source of Faunal Resemblances.—Effects of Direction of Shore Lines.—Numbers of Genera in Different Faunas.—Significance of Rare Forms.—Distribution of Shore-fishes.—Extension of Indian Fauna.—The Isthmus of Suez as a Barrier to Distribution.—Geological Evidences of Submergence of Isthmus of Suez.—The Cape of Good Hope as a Barrier to Fishes.—Relations of Japan to the Mediterranean Explained by Present Conditions.—The Isthmus of Panama as a Barrier to Distribution.—Unlikeness of Species on the Shores of the Isthmus of Panama.—Views of Dr. Günther on the Isthmus of Panama.—Catalogue of Fishes of Panama.—Conclusions of Evermann & Jenkins.—Conclusions of Dr. Hill.—Final Hypothesis as to Panama.
255CHAPTER XVI.
DISPERSION OF FRESH-WATER FISHES.
The Dispersion of Fishes.—The Problem of Oatka Creek.—Generalizations as to Dispersion.—Questions Raised by Agassiz.—Conclusions of Cope.—Questions Raised by Cope.—Views of Günther.—Fresh-water Fishes of North America.—Characters of Species.—Meaning of Species.—Special Creation Impossible.—Origin of American Species of Fishes.
282CHAPTER XVII.
DISPERSION OF FRESH-WATER FISHES. (
Continued.)
Barriers to Dispersion of Fresh-water Fishes: Local Barriers.—Favorable Waters Have Most Species.—Watersheds.—How Fishes Cross Watersheds.—The Suletind.—The Cassiquiare.—Two-Ocean Pass.—Mountain Chains.—Upland Fishes.—Lowland Fishes.—Cuban Fishes.—Swampy Watersheds.—The Great Basin of Utah.—Arctic Species in Lakes.—Causes of Dispersion still in Operation.
297CHAPTER XVIII.
FISHES AS FOOD FOR MAN.
The Flesh of Fishes.—Relative Rank of Food-fishes.—Abundance of Food-fishes.—Variety of Tropical Fishes.—Economic Fisheries.—Angling.
320CHAPTER XIX.
DISEASES OF FISHES.
Contagious Diseases: Crustacean Parasites.—Myxosporidia or Parasitic Protozoa.—Parasitic Worms: Trematodes, Cestodes.—The Worm of the Yellowstone.—The Heart Lake Tape-worm.—Thorn-head Worms.—Nematodes.—Parasitic Fungi.—Earthquakes.—Mortality of Filefish.
340CHAPTER XX.
THE MYTHOLOGY OF FISHES.
The Mermaid.—The Monkfish.—The Bishop-fish.—The Sea-serpent.
359CHAPTER XXI.
THE CLASSIFICATION OF FISHES.
Taxonomy.—Defects in Taxonomy.—Analogy and Homology.—Coues on Classification.—Species as Twigs of a Genealogical Tree.—Nomenclature.—The Conception of Genus and Species.—The Trunkfishes.—Trinomial Nomenclature.—Meaning of Species.—Generalization and Specialization.—High and Low Forms.—The Problem of the Highest Fishes.
367CHAPTER XXII.
THE HISTORY OF ICHTHYOLOGY.
Aristotle.—Rondelet.—Marcgraf.—Osbeck.—Artedi.—Linnæus.— Forskål.—Risso.—Bloch.—Lacépède.—Cuvier.—Valenciennes.— Agassiz.—Bonaparte.—Günther.—Boulenger.—Le Sueur.—Müller.— Gill.—Cope.—Lütken.—Steindachner.—Vaillant.—Bleeker.— Schlegel.—Poey.—Day.—Baird.—Garman.—Gilbert.—Evermann.— Eigenmann.—Zittel.—Traquair.—Woodward.—Dean.—Eastman.—Hay.— Gegenbaur.—Balfour.—Parker.—Dollo.
387CHAPTER XXIII.
THE COLLECTION OF FISHES.
How to Secure Fishes.—How to Preserve Fishes.—Value of Formalin.—Records of Fishes.—Eternal Vigilance.
429CHAPTER XXIV.
THE EVOLUTION OF FISHES.
The Geological Distribution of Fishes.—The Earliest Sharks.—Devonian Fishes.—Carboniferous Fishes.—Mesozoic Fishes.—Tertiary Fishes.—Factors of Extinction.—Fossilization of a Fish.—The Earliest Fishes.—The Cyclostomes.—The Ostracophores.—The Arthrodires.—The Sharks.—Origin of the Shark.—The Chimæras.—The Dipnoans.—The Crossopterygians.—The Actinopteri.—The Bony Fishes.
435CHAPTER XXV.
THE PROTOCHORDATA.
The Chordate Animals.—The Protochordates.—Other Terms Used in Classification.—The Enteropneusta.—Classification of Enteropneusta.—Family Harrimaniidæ.—Balanoglossidæ.—Low Organization of Harrimaniidæ.
460CHAPTER XXVI.
THE TUNICATES, OR ASCIDIANS.
Structure of Tunicates.—Development of Tunicates.—Reproduction of Tunicates.—Habits of Tunicates.—Larvacea.—Ascidiacea.—Thaliacea.—Origin of Tunicates.—Degeneration of Tunicates.
467CHAPTER XXVII.
THE LEPTOCARDII, OR LANCELETS.
The Lancelet.—Habits of Lancelets.—Species of Lancelets.—Origin of Lancelets.
482CHAPTER XXVIII.
THE CYCLOSTOMES, OR LAMPREYS.
The Lampreys.—Structure of the Lamprey.—Supposed Extinct Cyclostomes.—Conodontes.—Orders of Cyclostomes.—The Hyperotreta, or Hagfishes.—The Hyperoartia, or Lampreys.—Food of Lampreys.—Metamorphosis of Lampreys.—Mischief Done by Lampreys.—Migration or "Running" of Lampreys.—Requisite Conditions for Spawning with Lampreys.—The Spawning Process with Lampreys.—What Becomes of Lampreys after Spawning?
486CHAPTER XXIX.
THE CLASS ELASMOBRANCHII, OR SHARK-LIKE FISHES.
The Sharks.—Characters of Elasmobranchs.—Classification of Elasmobranchs.—Subclasses of Elasmobranchs.—The Selachii.—Hasse's Classification of Elasmobranchs.—Other Classifications of Elasmobranchs.—Primitive Sharks.—Order Pleuropterygii.—Order Acanthodii.—Dean on Acanthodii.—Order Ichthyotomi.
506CHAPTER XXX.
THE TRUE SHARKS.
Order Notidani.—Family Hexanchidæ.—Family Chlamydoselachidæ.—Order Asterospondyli.—Suborder Cestraciontes.—Family Heterodontidæ.—Edestus and its Allies.—Onchus.—Family Cochliodontidæ.—Suborder Galei.—Family Scyliorhinidæ.—The Lamnoid, or Mackerel-sharks.—Family Mitsukurinidæ, the Goblin-sharks.—Family Alopiidæ, or Thresher-sharks.—Family Pseudotriakidæ.—Family Lamnidæ.—Man-eating Sharks.—Family Cetorhinidæ, or Basking Sharks.—Family Rhineodontidæ.—The Carcharioid Sharks, or Requins.—Family Sphyrnidæ, or Hammer-head Sharks.—The Order of Tectospondyli.—Suborder Cyclospondyli.—Family Squalidæ.—Family Dalatiidæ.—Family Echinorhinidæ.—Suborder Rhinæ.—Family Pristiophoridæ, or Saw-sharks.—Suborder Batoidei, or Rays.—Pristididæ, or Sawfishes.—Rhinobatidæ, or Guitar-fishes.—Rajidæ, or Skates.—Narcobatidæ, or Torpedoes.—Petalodontidæ.—Dasyatidæ, or Sting-rays.—Myliobatidæ.—Family Psammodontidæ.—Family Mobulidæ. 523
CHAPTER XXXI.
THE HOLOCEPHALI, OR CHIMÆRAS.
The Chimæras.—Relationship of Chimæras.—Family Chimæridæ.—Rhinochimæridæ.—Extinct Chimæroids.—Ichthyodorulites.
561CHAPTER XXXII.
THE CLASS OSTRACOPHORI.
Ostracophores.—Nature of Ostracophores.—Orders of Ostracophores.—Order Heterostraci.—Order Osteostraci.—Order Antiarcha.—Order Anaspida.
568CHAPTER XXXIII.
ARTHRODIRES.
The Arthrodires.—Occurrence of Arthrodires.—Arthrognathi.—Anarthrodira.—Stegothalami.— Arthrodira.—Temnothoraci.—Arthrothoraci.—Relations of
Arthrodires.—Suborder Cycliæ.—Palæospondylus.—Gill on Palæospondylus.—Views as to the Relationships of Palæospondylus: Huxley, Traquair, 1890. Traquair, 1893. Traquair, 1897. Smith Woodward, 1892. Dawson, 1893. Gill, 1896. Dean, 1896. Dean, 1898. Parker & Haswell, 1897. Gegenbaur, 1898.—Relationships of Palæospondylus
581CHAPTER XXXIV.
THE CROSSOPTERYGII.
Class Teleostomi.—Subclass Crossopterygii.—Order of Amphibians.—The Fins of Crossopterygians.—Orders of Crossopterygians.—Haplistia.—Rhipidistia.—Megalichthyidæ.—Order Actinistia.—Order Cladistia.—The Polypteridæ
598CHAPTER XXXV.
SUBCLASS DIPNEUSTI, OR LUNGFISHES.
The Lungfishes.—Classification of Dipnoans.—Order Ctenodipterini.—Order Sirenoidei.—Family Ceratodontidæ.—Development of Neoceratodus.—Lepidosirenidæ.—Kerr on the Habits of Lepidosiren
609FOOTNOTES:
[1] For most of this list of errata I am indebted to the kindly interest of Dr. B. W. Evermann.
LIST OF ILLUSTRATIONS
VOL. I.
mesocoracoid
Some years since, at Madras, I (Dr. Day) obtained several specimens of a fresh-water Siluroid fish (Macrones vittatus) which is termed the "fiddler" in Mysore. I touched one which was on the wet ground, at which it appeared to become very irate, erecting its dorsal fin, making a noise resembling the buzzing of a bee. Having put some small carp into an aquarium containing one of these fishes, it rushed at a small example, seized it by the middle of its back, and shook it like a dog killing a rat; at this time its barbels were stiffened out laterally like a cat's whiskers.
It is noteworthy that the upland fishes are nearly the same in all these streams until we reach the southern limit of possible glacial influence. South of western North Carolina the faunæ of the different river basins appear to be more distinct from one another. Certain ripple-loving types are represented by closely related but unquestionably different species in each river basin, and it would appear that a thorough mingling of the upland species in these rivers has never taken place.
Very near the first among sea-fishes must come the pampano (Trachinotus carolinus) of the Gulf of Mexico, with firm, white, finely flavored flesh.
The various kinds of trout have been made famous the world over. All are attractive in form and color; all are gamey; all have the most charming of scenic surroundings, and, finally, all are excellent as food, not in the first rank perhaps, but well above the second. Notable among these are the European charr (Salvelinus alpinus), the American speckled trout or charr (Salvelinus fontinalis), the Dolly Varden or malma (Salvelinus malma), and the oquassa trout (Salvelinus oquassa). Scarcely less attractive are the true trout, the brown trout, or forelle (Salmo fario), in Europe, the rainbow-trout (Salmo irideus), the steelhead (Salmo gairdneri), the cut-throat trout (Salmo clarkii), and the Tahoe trout (Salmo henshawi), in America, and the yamabe (Salmo perryi) of Japan. Not least of all these is the flower of fishes, the grayling (Thymallus), of different species in different parts of the world.
Earthquakes.—Occasionally an earthquake has been known to kill sea-fishes in large numbers. The Albatross obtained specimens of Sternoptyx diaphana in the Japanese Kuro Shiwo, killed by the earthquakes of 1896, which destroyed fishing villages of the coast of Rikuchu in northern Japan.
A perfect taxonomy is one which would perfectly express all the facts in the evolution and development of the various forms. It would recognize all the evidence from the three ancestral documents, palæontology, morphology, and ontogeny. It would consider structure and form independently of adaptive or physiological or environmental modifications. It would regard as most important those characters which had existed longest unchanged in the history of the species or type. It would regard as of first rank those characters which appear first in the history of the embryo. It would regard as of minor importance those which had arisen recently in response to natural selection or the forced alteration through pressure of environment, while fundamental alterations as they appear one after another in geologic time would make the basal characters of corresponding groups in taxonomy. In a perfect taxonomy or natural system of classification animals would not be divided into groups nor ranged in linear series. We should imagine series variously and divergently branched, with each group at its earlier or lower end passing insensibly into the main or primitive stock. A very little alteration now and then in some structure is epoch-making, and paves the way through specialization to a new class or order. But each class or order through its lowest types is interlocked with some earlier and otherwise diverging group.
The scanty pre-Cuvieran work on the fishes of North America has been already noticed. Contemporary with the early work of Cuvier is the worthy attempt of Professor Samuel Latham Mitchill (1764-1831) to record in systematic fashion the fishes of New York. Soon after followed the admirable work of Charles Alexandre Le Sueur (1789-1840), artist and naturalist, who was the first to study the fishes of the Great Lakes and the basin of the Ohio. Le Sueur's engravings of fishes, in the early publications of the Academy of Natural Sciences in Philadelphia, are still among the most satisfactory representations of the species to which they refer. Constantine Samuel Rafinesque (1784-1842), the third of this remarkable but very dissimilar trio, published numerous papers descriptive of the species he had seen or heard of in his various botanical rambles. This culminated in his elaborate but untrustworthy "Ichthyologia Ohiensis." The fishes of Ohio received later a far more conscientious though less brilliant treatment at the hands of Dr. Jared Potter Kirtland (1793-1877), an eminent physician of Cleveland, Ohio. In 1842 the amiable and scholarly James Ellsworth Dekay (1799-1851) published his detailed report on the fishes of the "New York Fauna," and a little earlier (1836) in the "Fauna Boreali-Americana" Sir John Richardson (1787-1865) gave a most valuable and accurate account of the fishes of the Great Lakes and Canada. Almost simultaneously, Rev. Zadock Thompson (1796-1856) gave a catalogue of the fishes of Vermont, and David Humphreys Storer (1804-91) began his work on the fishes of Massachusetts, finally expanded into a "Synopsis of the Fishes of North America" (1846) and a "History of the Fishes of Massachusetts" (1853-67). Dr. John Edwards Holbrook (1794-1871), of Charleston, published (1855-60) his invaluable record of the fishes of South Carolina, the promise of still more important work, which was prevented by the outbreak of the Civil War in the United States. The monograph on Lake Superior (1850) and other publications of Louis Agassiz (1807-73) have been already noticed. One of the first of Agassiz's students was Charles Girard (1822-95), who came with him from Switzerland, and, in association with Spencer Fullerton Baird (1823-87), described the fishes from the United States Pacific Railway Surveys (1858) and the United States and Mexican Boundary Surveys (1859). Professor Baird, primarily an ornithologist, became occupied with executive matters, leaving Girard to finish these studies of the fishes. A large part of the work on fishes published by the United States National Museum and the United States Fish Commission has been made possible through the direct help and inspiration of Professor Baird. Among those engaged in this work, James William Milner (1841-80), Marshall Macdonald (1836-95), and Hugh M. Smith may be noted.
Other faunal writers of more or less prominence were William Dandridge Peck (1763-1822) in New Hampshire, George Suckley (1830-69) in Oregon, James William Milner (1841-80) in the Great Lake Region, Samuel Stehman Haldeman (1812-80) in Pennsylvania, William O. Ayres (1817-91) in Connecticut and California; Dr. John G. Cooper (died 1902), Dr. William P. Gibbons and Dr. William N. Lockington (died 1902) in California; Philo Romayne Hoy (1816-93) studied the fishes of Wisconsin, Charles Conrad Abbott those of New Jersey, Silas Stearns (1859-88) those of Florida, Stephen Alfred Forbes and Edward W. Nelson those of Illinois, Oliver Perry Hay, later known for his work on fossil forms, those of Mississippi, Alfredo Dugés, of Guanajuato, those of Central Mexico.
Third Period.—Morphological Work on Fossil Fishes.—Among the writers who have dealt with the problems of the relationships of the Ostracophores as well as Palæospondylus and the Arthrodires may be named Traquair, Huxley, Newberry, Smith Woodward, Rohon, Eastman, and Dean; most recently William Patten. Upon the phylogeny of the sharks Traquair, A. Fritsch, Hasse, Cope, Brongniart, Jaekel, Reis, Eastman, and Dean. On Chimæroid morphology mention may be made of the papers of A. S. Woodward, Reis, Jaekel, Eastman, C. D. Walcott, and Dean. As to Dipnoan relationships the paper of Louis Dollo is easily of the first value; of especial interest, too, is the work of Eastman as to the early derivation of the Dipnoan dentition. In this regard a paper of Rohon is noteworthy, as is also that of Richard Semon on the development of the dentition of recent Neoceratodus, since it contains a number of references to extinct types. Interest notes on Dipnoan fin characters have been given by Traquair. In the morphology of Ganoids, the work of Traquair and A. S. Woodward takes easily the foremost rank. Other important works are those of Huxley, Cope, A. Fritsch, and Oliver P. Hay.
The known branchiferous or gill-bearing chordates living and extinct may be first divided into eight classes—the Enteropneusta, the Tunicata, the Leptocardii, or lancelets, the Cyclostomi, or lampreys, the Elasmobranchii, or sharks, the Ostracophori the Arthrodira, and the Teleostomi, or true fishes. The first two groups, being very primitive and in no respect fish-like in appearance, are sometimes grouped together as Protochordata, the others with the higher Chordates constituting the Vertebrata.
[Pg 367]
ERRATA[1]
VOL. I
PAGE
Lepomis megalotis, Long-eared Sunfish
2 Lepomis megalotis, Long-eared Sunfish
4 Eupomotis gibbosus, Common Sunfish
7 Ozorthe dictyogramma, a Japanese Blenny
9 Eupomotis gibbosus, Common Sunfish
13 Monocentris japonicas, Pine-cone Fish
16 Diodon hystrix, Porcupine-fish
17 Nemichthys avocetta, Thread-eel
17 Hippocampus hudsonius, Sea-horse
17 Peprilus paru, Harvest-fish
18 Lophius litulon, Anko or Fishing-frog
18 Epinephelus adscensionis, Rock-hind or Cabra Mora
20Scales of
Acanthoessus bronni 21Cycloid Scale
22 Porichthys porosissimus, Singing-fish
23 Apomotis cyanellus, Blue-green Sunfish
27 Chiasmodon niger, Black Swallower
29Jaws of a Parrot-fish,
Sparisoma aurofrenatum 30 Archosargus probatocephalus, Sheepshead
31 Campostoma anomalum, Stone-roller
33 Roccus lineatus, Striped Bass
35 Roccus lineatus.Lateral View of Cranium
36 Roccus lineatus.Superior View of Cranium
37 Roccus lineatus.Inferior View of Cranium
38 Roccus lineatus. Posterior View of Cranium
40 Roccus lineatus.Face-bones, Shoulder and Pelvic Girdles, and Hyoid Arch
42Lower Jaw of
Amia calva, showing Gular Plate
43 Roccus lineatus.Branchial Arches
46Pharyngeal Bone and Teeth of European Chub,
Leuciscus cephalus 47Upper Pharyngeals of Parrot-fish,
Scarus strongylocephalus 47Lower Pharyngeal Teeth of Parrot-fish,
Scarus strongylocephalus 47Pharyngeals of Italian Parrot-fish,
Sparisoma cretense 48 Roccus lineatus, Vertebral Column and Appendages
48Basal Bone of Dorsal Fin,
Holoptychius leptopterus 49Inner View of Shoulder-girdle of Buffalo-fish,
Ictiobus bubalus 51 Pterophryne tumida, Sargassum-fish.
52Shoulder-girdle of
Sebastolobus alascanus.
52Cranium of
Sebastolobus alascanus.
53Lower Jaw and Palate of
Sebastolobus alascanus.
54Maxillary and Premaxillary of
Sebastolobus alascanus.
55Part of Skeleton of
Selene vomer.
55Hyostylic Skull of
Chiloscyllium indicum, a Scyliorhinoid Shark.
56Skull of
Heptranchias indicus, a Notidanoid Shark.
56Basal Bones of Pectoral Fin of Monkfish,
Squatina.
56Pectoral Fin of
Heterodontus philippi.
57Pectoral Fin of
Heptranchias indicus.
57Shoulder-girdle of a Flounder,
Paralichthys californicus.
58Shoulder-girdle of a Toadfish,
Batrachoides pacifici.
59Shoulder-girdle of a Garfish,
Tylosurus fodiator.
59Shoulder-girdle of a Hake,
Merluccius productus.
60 Cladoselache fyleri, Restored.
65Fold-like Pectoral and Ventral Fins of
Cladoselache fyleri.
65Pectoral Fin of a Shark,
Chiloscyllium.
66Skull and Shoulder-girdle of
Neoceratodus forsteri, showing archipterygium.
68 Acanthoessus wardi.
69Shoulder-girdle of
Acanthoessus.
69Pectoral Fin of
Pleuracanthus.
69Shoulder-girdle of
Polypterus bichir.
70Arm of a Frog.
71 Pleuracanthus decheni.
74Embryos of
Heterodontus japonicas, a Cestraciont Shark.
75 Polypterus congicus, a Crossopterygian Fish with External Gills.
78Heterocercal Tail of Sturgeon,
Acipenser sturio.
80Heterocercal Tail of Bowfin,
Amia calva.
82Heterocercal Tail of Garpike,
Lepisosteus osseus.
82 Coryphænoides carapinus, showing Leptocercal Tail.
83Heterocercal Tail of Young Trout,
Salmo fario.
83Isocercal Tail of Hake,
Merluccius productus.
84Homocercal Tail of a Flounder,
Paralichthys californicus.
84Gephyrocercal Tail of
Mola mola.
85Shoulder-girdle of
Amia calva.
86Shoulder-girdle of a Sea-catfish,
Selenaspis dowi.
86Clavicles of a Sea-catfish,
Selenaspis dowi.
87Shoulder-girdle of a Batfish,
Ogcocephalus radiatus.
88Shoulder-girdle of a Threadfin,
Polydactylus approximans.
89Gill-basket of Lamprey.
92Weberian Apparatus and Air-bladder of Carp.
93Brain of a Shark,
Squatina squatina.
110Brain of
Chimæra monstrosa.
110Brain of
Polypterus annectens.
110Brain of a Perch,
Perca flavescens.
111 Petromyzon marinus unicolor.Head of Lake Lamprey, showing Pineal Body.
111 Chologaster cornutus, Dismal-swamp Fish.
116 Typhlichthys subterraneus, Blind Cavefish.
116 Anableps dovii, Four-eyed Fish.
117Ipnops murrayi.
118 Boleophthalmus chinensis, Pond-skipper.
118 Lampetra wilderi, Brook Lamprey.
120 Branchiostoma lanceolatum, European Lancelet.
120 Pseudupeneus maculatus, Goatfish.
122 Xiphophorus helleri, Sword-tail Minnow.
124 Cymatogaster aggregatus, White Surf-fish, Viviparous, with Young.
125 Goodea luitpoldi, a Viviparous Fish.
126Egg of
Callorhynchus antarcticus, the Bottle-nosed Chimæra.
127Egg of the Hagfish,
Myxine limosa.
127Egg of Port Jackson Shark,
Heterodontus philippi.
128Development of Sea-bass,
Centropristes striatus.
135 Centropristes striatus, Sea-bass.
137 Xiphias gladius, Young Sword-fish.
139 Xiphias gladius, Sword-fish.
139Larva of the Sail-fish,
Istiophorus, Very Young.
140Larva of Brook Lamprey,
Lampetra wilderi, before Transformation.
140 Anguilla chrisypa, Common Eel.
140Larva of Common Eel,
Anguilla chrisypa, called
Leptocephalus grassii.
141Larva of Sturgeon,
Acipenser sturio.
141Larva of
Chætodon sedentarius.
142 Chætodon capistratus, Butterfly-fish.
142 Mola mola, Very Early Larval Stage of Headfish, called
Centaurus boöps.
143 Mola mola, Early Larval Stage called
Molacanthus nummularis.
144 Mola mola, Advanced Larval Stage.
144 Mola mola, Headfish, Adult.
146 Albula vulpes, Transformation of Ladyfish from Larva to Young.
147Development of the Horsehead-fish,
Selene vomer.
148 Salanx hyalocranius, Ice-fish.
149 Dallia pectoralis, Alaska Blackfish.
149 Ophiocephalus barca, Snake-headed China-fish.
150 Carassius auratus, Monstrous Goldfish.
151Jaws of
Nemichthys avocetta.
156 Cypsilurus californicus, Flying-fish.
157 Ammocrypta clara, Sand-darter.
158 Fierasfer acus, Pearl-fish, issuing from a Holothurian.
159 Gobiomorus gronovii, Portuguese Man-of-war Fish.
160Tide Pools of Misaki.
161 Ptychocheilus oregonensis, Squaw-fish.
162 Ptychocheilus grandis, Squaw-fish, Stranded as the Water Falls.
164Larval Stages of
Platophrys podas, a Flounder of the Mediterranean, showing Migration of Eye.
174 Platophrys lunatus, the Wide-eyed Flounder.
175Young Flounder Just Hatched, with Symmetrical Eyes.
175 Pseudopleuronectes americanus, Larval Flounder.
176 Pseudopleuronectes americanus, Larval Flounder (more advanced stage).
176Face View of Recently-hatched Flounder.
177 Schilbeodes furiosus, Mad-Tom.
179 Emmydrichthys vulcanus, Black Nohu or Poison-fish.
180 Teuthis bahianus, Brown Tang.
181 Stephanolepis hispidus, Common Filefish.
182Tetraodon meleagris.
183 Balistes carolinensis, the Trigger-fish.
184 Narcine brasiliensis, Numbfish.
185 Torpedo electricus, Electric Catfish.
186 Astroscopus guttatus, Star-gazer.
187 Æthoprora lucida, Headlight-fish.
188 Corynolophus reinhardti, showing Luminous Bulb.
188Etmopterus lucifer.
189Argyropelecus olfersi.
190Luminous Organs and Lateral Line of Midshipman,
Porichthys notatus.
192Cross-section of Ventral Phosphorescent Organ of Midshipman,
Porichthys notatus.
193Section of Deeper Portion of Phosphorescent Organ,
Porichthys notatus.
194 Leptecheneis naucrates, Sucking-fish or Pegador.
197 Caularchus mæandricus, Clingfish.
198 Polistotrema stouti, Hagfish.
199 Pristis zysron, Indian Sawfish.
200 Pristiophorus japonicus, Saw-shark.
201Skeleton of Pike,
Esox lucius.
203Skeleton of Red Rockfish,
Sebastodes miniatus.
214Skeleton of a Spiny-rayed Fish of the Tropics,
Holacanthus ciliaris.
214Skeleton of the Cowfish,
Lactophrys tricornis.
215 Crystallias matsushimæ, Liparid.
218 Sebastichthys maliger, Yellow-backed Rockfish.
218 Myoxocephalus scorpius, European Sculpin.
219 Hemitripterus americanus, Sea-raven.
220 Cyclopterus lumpus, Lumpfish.
220 Psychrolutes paradoxus, Sleek Sculpin.
221 Pallasina barbata, Agonoid-fish.
221 Amblyopsis spelæus, Blindfish of the Mammoth Cave.
221 Lucifuga subterranea, Blind Brotula.
222 Hypsypops rubicunda, Garibaldi.
227 Synanceia verrucosa, Gofu or Poison-fish.
229 Alticus saliens, Lizard-skipper.
230 Etheostoma camurum, Blue-breasted Darter.
231 Liuranus semicinctusand
Chlevastes colubrinus, Snake-eels.
233Coral Reef at Apia.
234 Rudarius ercodes, Japanese Filefish.
241 Tetraodon setosus, Globefish.
244 Dasyatis sabina, Sting-ray.
246 Diplesion blennioides, Green-sided Darter.
247 Hippocampus mohnikei, Japanese Sea-horse.
250 Archoplites interruptus, Sacramento Perch.
258Map of the Continents, Eocene Time.
270 Caulophryne jordani, Deep-sea Fish of Gulf Stream.
276 Exerpes asper, Fish of Rock-pools, Mexico.
276Xenocys jessiæ.
279 Ictalurus punctatus, Channel Catfish.
280Drawing the Net on the Beach of Hilo, Hawaii.
281 Semotilus atromaculatus, Horned Dace.
285 Leuciscus lineatus, Chub of the Great Basin.
287 Melletes papilio, Butterfly Sculpin.
288 Scartichthys enosimæ, a Fish of the Rock-pools of the Sacred Island of Enoshima, Japan.
294 Halichœres bivittatus, the Slippery Dick.
297Peristedion miniatum.
299Outlet of Lake Bonneville.
303 Hypocritichthys analis, Silver Surf-fish.
309 Erimyzon sucetta, Creekfish or Chub-sucker.
315 Thaleichthys pretiosus, Eulachon or Ulchen.
320 Plecoglossus altivelis, the Japanese Ayu.
321 Coregonus clupeiformis, the Whitefish.
321 Mullus auratus, the Golden Surmullet.
322 Scomberomorus maculatus, the Spanish Mackerel.
322 Lampris luna, the Opah or Moonfish.
323 Pomatomus saltatrix, the Bluefish.
324 Centropomus undecimalis, the Robalo.
324 Chætodipterus faber, the Spadefish.
325 Micropterus dolomieu, the Small-mouthed Black Bass.
325 Salvelinus fontinalis, the Speckled Trout.
326 Salmo irideus, the Rainbow Trout.
326 Salvelinus oquassa, the Rangeley Trout.
326 Salmo gairdneri, the Steelhead Trout.
327 Salmo henshawi, the Tahoe Trout.
327 Salvelinus malma, the Dolly Varden Trout.
327 Thymallus signifer, the Alaska Grayling.
328 Esox lucius, the Pike.
328 Pleurogrammus monopterygius, the Atka-fish.
328 Chirostoma humboldtianum, the Pescado blanco.
329 Pseudupeneus maculatus, the Red Goatfish.
329 Pseudoscarus guacamaia, Great Parrot-fish.
330 Mugil cephalus, Striped Mullet.
330 Lutianus analis, Mutton-snapper.
331 Clupea harengus, Herring.
331 Gadus callarias, Codfish.
331 Scomber scombrus, Mackerel.
332 Hippoglossus hippoglossus, Halibut.
332Fishing for Ayu with Cormorants.
333Fishing for Ayu. Emptying Pouch of Cormorant.
335Fishing for Tai, Tokyo Bay.
338 Brevoortia tyrannus, Menhaden.
340 Exonautes unicolor, Australian Flying-fish.
341 Rhinichthys atronasus, Black-nosed Dace.
342 Notropis hudsonius, White Shiner.
343 Ameiurus catus, White Catfish.
344 Catostomus ardens, Sucker.
348 Oncorhynchus tschawytscha, Quinnat Salmon.
354 Oncorhynchus tschawytscha, Young Male.
355 Ameiurus nebulosus, Catfishes.
358"Le Monstre Marin en Habit de Moine".
360"Le Monstre Marin en Habit d'Évêque".
361 Regalecus russelli, Oarfish.
362 Regalecus glesne, Glesnæs Oarfish.
363 Nemichthys avocetta, Thread-eel.
365 Lactophrys tricornis, Horned Trunkfish.
373 Ostracion cornutum, Horned Trunkfish.
376 Lactophrys bicaudalis, Spotted Trunkfish.
377 Lactophrys bicaudalis, Spotted Trunkfish (Face).
377 Lactophrys triqueter, Spineless Trunkfish.
378 Lactophrys trigonus, Hornless Trunkfish.
378 Lactophrys trigonus, Hornless Trunkfish (Face).
379Bernard Germain de Lacépède.
399Georges Dagobert Cuvier.
399Louis Agassiz.
399Johannes Müller.
399Albert Günther.
403Franz Steindachner.
403George Albert Boulenger.
403Robert Collett.
403Spencer Fullerton Baird.
407Edward Drinker Cope.
407Theodore Nicholas Gill.
407George Brown Goode.
407Johann Reinhardt.
409Edward Waller Claypole.
409Carlos Berg.
409Edgar R. Waite.
409Felipe Poey y Aloy.
413Léon Vaillant.
413Louis Dollo.
413Decio Vinciguerra.
413Bashford Dean.
417Kakichi Mitsukuri.
417Carl H. Eigenmann.
417Franz Hilgendorf.
417David Starr Jordan.
421Herbert Edson Copeland.
421Charles Henry Gilbert.
421Barton Warren Evermann.
421Ramsay Heatley Traquair.
425Arthur Smith Woodward.
425Karl A. Zittel.
425Charles R. Eastman.
425Fragment of Sandstone from Ordovician Deposits.
435Fossil Fish Remains from Ordovician Rocks.
436Dipterus valenciennesi.
437Hoplopteryx lewesiensis.
438 Paratrachichthys prosthemius, Berycoid fish.
439 Cypsilurus heterurus, Flying-fish.
440 Lutianidæ, Schoolmaster Snapper.
440 Pleuronichthys decurrens, Decurrent Flounder.
441 Cephalaspis lyelli, Ostracophore.
444 Dinichthys intermedius, Arthrodire.
445 Lamna cornubica, Mackerel-shark or Salmon-shark.
447 Raja stellulata, Star-spined Ray.
448 Harriotta raleighiana, Deep-sea Chimæra.
449 Dipterus valenciennesi, Extinct Dipnoan.
449 Holoptychius giganteus, Extinct Crossopterygian.
451 Platysomus gibbosus, Ancient Ganoid fish.
452 Lepisosteus platystomus, Short-nosed Gar.
452 Palæoniscum macropomum, Primitive Ganoid fish.
453 Diplomystus humilis, Fossil Herring.
453 Holcolepis lewesiensis.
454 Elops saurus, Ten-pounder.
454 Apogon semilineatus, Cardinal-fish.
455 Pomolobus æstivalis, Summer Herring.
455Bassozetus catena.
456Trachicephalus uranoscopus.
456 Chlarias breviceps, African Catfish.
457 Notropis whipplii, Silverfin.
457Gymnothorax moringa.
458 Seriola lalandi, Amber-fish.
458Geological Distribution of the Families of Elasmobranchs.
459"Tornaria" Larva of
Glossobalanus minutus.
463Glossobalanus minutus.
464Harrimania maculosa.
465Development of Larval Tunicate to Fixed Condition.
471Anatomy of Tunicate.
472Ascidia adhærens.
474Styela yacutatensis.
475Styela greeleyi.
476Cynthia superba.
476 Botryllus magnus, Compound Ascidian.
477Botryllus magnus.
478 Botryllus magnus, a Single Zooid.
479 Aplidiopsis jordani, a Compound Ascidian.
479 Oikopleura, Adult Tunicate of Group Larvacea.
480 Branchiostoma californiense, California Lancelet.
484Gill-basket of Lamprey.
485Polygnathus dubium.
488 Polistotrema stouti, Hagfish.
489 Petromyzon marinus, Lamprey.
491 Petromyzon marinus unicolor, Mouth Lake Lamprey.
492 Lampetra wilderi, Sea Larvæ Brook Lamprey.
492 Lampetra wilderi, Mouth Brook Lamprey.
492 Lampetra camtschatica, Kamchatka Lamprey.
495 Entosphenus tridentatus, Oregon Lamprey.
496 Lampetra wilderi, Brook Lamprey.
505Fin-spine of
Onchus tenuistriatus.
509Section of Vertebræ of Sharks, showing Calcification.
510Cladoselache fyleri.
514 Cladoselache fyleri, Ventral View.
515Teeth of
Cladoselache fyleri.
515Acanthoessus wardi.
515Diplacanthus crassissimus.
517Climatius scutiger.
518Pleuracanthus decheni.
519 Pleuracanthus decheni, Restored.
520Head-bones and Teeth of
Pleuracanthus decheni.
520Teeth of
Didymodus bohemicus.
520Shoulder-girdle and Pectoral Fins of
Cladodus neilsoni.
521Teeth of
Cladodus striatus.
522 Hexanchus griseus, Griset or Cow-shark.
523Teeth of
Heptranchias indicus.
524 Chlamydoselachus anguineus, Frill-shark.
525 Heterodontus francisci, Bullhead-shark.
526Lower Jaw of
Heterodontus philippi.
526Teeth of Cestraciont Sharks.
527Egg of Port Jackson Shark,
Heterodontus philippi.
527Tooth of
Hybodus delabechei.
528Fin-spine of
Hybodus basanus.
528Fin-spine of
Hybodus reticulatus.
528Fin-spine of
Hybodus canaliculatus.
529Teeth of Cestraciont Sharks.
529 Edestus vorax, Supposed to be a Whorl of Teeth.
529 Helicoprion bessonowi, Teeth of.
530Lower Jaw of
Cochliodus contortus.
531 Mitsukurina owstoni, Goblin-shark.
535 Scapanorhynchus lewisi, Under Side of Snout.
536Tooth of
Lamna cuspidata.
537 Isuropsis dekayi, Mackerel-shark.
537Tooth of
Isurus hastalis.
538[Pg 2]
The Long-eared Sunfish.—If we would understand a fish, we must first go and catch one. This is not very hard to do, for there are plenty of them in the little rushing brook or among the lilies of the pond. Let us take a small hook, put on it an angleworm or a grasshopper,—no need to seek an elaborate artificial fly,—and we will go out to the old "swimming-hole" or the deep eddy at the root of the old stump where the stream has gnawed away the bank in changing its course. Here we will find fishes, and one of them will take the bait very soon. In one part of the country the first fish that bites will be different from the first one taken in some other. But as we are fishing in the United States, we will locate our brook in the centre of population of our country. This will be to the northwest of Cincinnati, among the low wooded hills from which clear brooks flow over gravelly bottoms toward the Ohio River. Here we will catch sunfishes of certain species, or maybe rock bass or catfish: any of these will do for our purpose. But one of our sunfishes is especially beautiful—mottled blue and golden and scarlet, with a long, black, ear-like appendage backward from his gill-covers—and this one we will keep and hold for our first lesson in fishes. It is a small fish, not longer than your hand most likely, but it can take the bait as savagely as the best, swimming away with it with such force that you might think from the vigor of its pull that you have a pickerel or a bass. But when it comes out of the water you see a little, flapping, unhappy, living plate of brown and blue and orange, with fins wide-spread and eyes red with rage.
Color of the Fish.—Behind the eye there are several bones on the side of the head which are more or less distinct from the skull itself. These are called membrane bones because they are formed of membrane which has become bony by the deposition in it of salts of lime. One of these is called the opercle, or gill-cover, and before it, forming a right angle, is the preopercle, or false gill-cover. On our sunfish we see that the opercle ends behind in a long and narrow flap, which looks like an ear. This is black in color, with an edging of scarlet as though a drop of blood had spread along its margin. When the fish is in the water its back is dark greenish-looking, like the weeds and the sticks in the bottom, so that we cannot see it very plainly. This is the way the fish looks to the fishhawks or herons in the air above it who may come to the stream to look for fish. Those fishes which from above look most like the bottom can most readily hide and save themselves. The under side of the sunfish is paler, and most fishes have the belly white. Fishes with white bellies swim high in the water, and the fishes who would catch them lie below. To the fish in the water all outside the water looks white, and so the white-bellied fishes are hard for other fishes to see, just as it is hard for us to see a white rabbit bounding over the snow.
When the sunfish is taken out of the water its colors seem to fade. In the aquarium it is generally paler, but it will sometimes brighten up when another of its own species is placed beside it. A cause of this may lie in the nervous control of the muscles at the base of the scales. When the scales lie very flat the color has one appearance. When they rise a little the shade of color seems to change. If you let fall some ink-drops between two panes of glass, then spread them apart or press them together, you will see changes in the color and size of the spots. Of this nature is the apparent change in the colors of fishes under different conditions. Where the fish feels at its best the colors are the richest. There are some fishes, too, in which the male grows very brilliant in the breeding season through the deposition of red, white, black, or blue pigments, or coloring matter, on its scales or on its head or fins, this pigment being absorbed when the mating season is over. This is not true of the sunfish, who remains just about the same at all seasons. The male and female are colored alike and are not to be distinguished without dissection. If we examine the scales, we shall find that these are marked with fine lines and concentric striæ, and part of the apparent color is due to the effect of the fine lines on the light. This gives the bluish lustre or sheen which we can see in certain lights, although we shall find no real blue pigment under it. The inner edge of each scale is usually scalloped or crinkled, and the outer margin of most of them has little prickly points which make the fish seem rough when we pass our hand along his sides.
The Brain of the Fish.—The movements of the fish, like those of every other complex animal, are directed by a central nervous system, of which the principal part is in the head and is known as the brain. From the eye of the fish a large nerve goes to the brain to report what is in sight. Other nerves go from the nostrils, the ears, the skin, and every part which has any sort of capacity for feeling. These nerves carry their messages inward, and when they reach the brain they may be transformed into movement. The brain sends back messages to the muscles, directing them to contract. Their contraction moves the fins, and the fish is shoved along through the water. To scare the fish or to attract it to its food or to its mate is about the whole range of the effect that sight or touch has on the animal. These sensations changed into movement constitute what is called reflex action, performance without thinking of what is being done. With a boy, many familiar actions may be equally reflex. The boy can also do many other things "of his own accord," that is, by conscious effort. He can choose among a great many possible actions. But a fish cannot. If he is scared, he must swim away, and he has no way to stop himself. If he is hungry, and most fishes are so all the time, he will spring at the bait. If he is thirsty, he will gasp, and there is nothing else for him to do. In other words, the activities of a fish are nearly all reflex, most of them being suggested and immediately directed by the influence of external things. Because its actions are all reflex the brain is very small, very primitive, and very simple, nothing more being needed for automatic movement. Small as the fish's skull-cavity is, the brain does not half fill it.
Thus, again, the length of the muzzle, the diameter of the eye, and other dimensions may be compared with the length of the head. In the Rock Hind, fig. 12, the eye is 5 in head, the snout is 4-2/5 in head, and the maxillary 2-3/5. Young fishes have the eye larger, the body slenderer, and the head larger in proportion than old fishes of the same kind. The mouth grows larger with age, and is sometimes larger also in the male sex. The development of the fins often varies a good deal in some fishes with age, old fishes and male fishes having higher fins when such differences exist. These variations are soon understood by the student of fishes and cause little doubt or confusion in the study of fishes.
Ctenoid and Cycloid Scales.—Normally formed scales are rounded in outline, marked by fine concentric rings, and crossed on the inner side by a few strong radiating ridges and folds. They usually cover the body more or less evenly and are imbricated like shingles on a roof, the free edge being turned backward. Such normal scales are of two types, ctenoid or cycloid. Ctenoid scales have a comb-edge of fine prickles or cilia; cycloid scales have the edges smooth. These two types are not very different, and the one readily passes into the other, both being sometimes seen on different parts of the same fish. In general, however, the more primitive representatives of the typical fishes, those with abdominal ventrals and without spines in the fins, have cycloid or smooth scales. Examples are the salmon, herring, minnow, and carp. Some of the more specialized spiny-rayed fishes, as the parrot-fishes, have, however, scales equally smooth, although somewhat different in structure. Sometimes, as in the eel, the cycloid scales may be reduced to mere rudiments buried in the skin.
Bony and Prickly Scales.—Bony and prickly scales are found in great variety, and scarcely admit of description or classification. In general, prickly points on the skin are modifications of ctenoid scales. Ganoid scales are thickened and covered with bony enamel, much like that seen in teeth, otherwise essentially like cycloid scales. These are found in the garpike and in many genera of extinct Ganoid and Crossopterygian fishes. In the line of descent the placoid scale preceded the ganoid, which in turn was followed by the cycloid and lastly by the ctenoid scale. Bony scales in other types of fishes may have nothing structurally in common with ganoid scales or plates, however great may be the superficial resemblance.
The Lateral Line as a Mucous Channel.—The more primitive condition of the lateral line is seen in the sharks and chimæras, in which fishes it appears as a series of channels in or under the skin. These channels are filled with mucus, which exudes through occasional open pores. In many fishes the bones of the skull are cavernous, that is, provided with cavities filled with mucus. Analogous to these cavities are the mucous channels which in primitive fishes constitute the lateral line.
The lancelets, lampreys, and hagfishes possess no genital ducts. In the former the germ cells are shed into the atrial cavity, and from there find their way to the exterior either through the mouth or the atrial pore; in the latter they are shed directly into the body cavity, from which they escape through the abdominal pores. In the sharks and skates the Wolffian duct in the male, in addition to its function as an excretory duct, serves also as a passage for the sperm, the testes having a direct connection with the kidneys. In these forms there is a pair of Müllerian ducts which serve as oviducts in the females; they extend the length of the body cavity, and at their anterior end have an opening which receives the eggs which have escaped from the ovary into the body cavity. In some bony fishes as the eels and female salmon the germ cells are shed into the body cavity and escape through genital pores, which, however, may not be homologous with abdominal pores. In most other bony fishes the testes and ovaries are continued directly into ducts which open to the outside.
The most usual type of teeth among fishes is that of villiform bands. Villiform teeth are short, slender, even, close-set, making a rough velvety surface. When the teeth are larger and more widely separated, they are called cardiform, like the teeth of a wool-card. Granular teeth are small, blunt, and sand-like. Canine teeth are those projecting above the level of the others, usually sharp, curved, and in some species barbed. Sometimes the canines are in front. In some families the last tooth in either jaw may be a "posterior canine," serving to hold small animals in place while the anterior teeth crush them. Canine teeth are often depressible, having a hinge at base.
Teeth very slender and brush-like are called setiform. Teeth with blunt tips are molar. These are usually enlarged and fitted for crushing shells. Flat teeth set in mosaic, as in many rays and in the pharyngeals of parrot-fishes, are said to be paved or tessellated. Knife-like teeth, occasionally with serrated edges, are found in many sharks. Many fishes have incisor-like teeth, some flattened and truncate like human teeth, as in the sheepshead, sometimes with serrated edges. Often these teeth are movable, implanted only in the skin of the lips. In other cases they are set fast in the jaw. Most species with movable teeth or teeth with serrated edges are herbivorous, while strong incisors may indicate the choice of snails and crabs as food. Two or more of these different types may be found in the same fish. The knife-like teeth of the sharks are progressively shed, new ones being constantly formed on the inner margins of the jaw, so that the teeth are marching to be lost over the edge of the jaw as soon as each has fulfilled its function. In general the more distinctly a species is a fish-eater, the sharper are the teeth. Usually fishes show little discrimination in their choice of food; often they devour the young of their own species as readily as any other. The digestive process is rapid, and most fishes rapidly increase in size in the process of development. When food ceases to be abundant the fishes grow more slowly. For this reason the same species will grow to a larger size in large streams than in small ones, in lakes than in brooks. In most cases there is no absolute limit to growth, the species growing as long as it lives. But while some species endure many years, others are certainly very shortlived, and some may be even annual, dying after spawning, perhaps at the end of the first season.
Lepisosteus
The assumption that each element in the shoulder-girdle and the pectoral fin of the fish must correspond in detail to the arm of man has led to great confusion in naming the different bones. Among the many bones of the fish's shoulder-girdle and pectoral fin, three or four different ones have successively borne the names of scapula, clavicle, coracoid, humerus, radius, and ulna. None of these terms, unless it be clavicle, ought by rights apply to the fish, for no bone of the fish is a true homologue of any of these as seen in man. The land vertebrates and the fishes have doubtless sprung from a common stock, but this stock, related to the crossopterygians of the present day, was unspecialized in the details of its skeleton, and from it the fishes and the higher vertebrates have developed the widely diverging lines.
In the sharks, dipnoans, crossopterygians, ganoids, and teleosts or bony fishes, jaws are developed as well as a variety of other bones around the mouth and throat. The jaw-bearing forms are sometimes known by the general name of gnathostomes. In the sharks and their relatives (rays, chimæras, etc.) all the skeleton is composed of cartilage. In the more specialized bony fishes, besides these bones we find also series of membrane bones, more or less external to the skull and composed of ossified dermal tissues. Membrane bones are not found in the sharks and lampreys, but are developed in an elaborate coat of mail in some extinct forms.
basisphenoid
The gill-arches are suspended to the cranium from above by the suspensory pharyngeal (44). Each arch contains three parts—the epibranchial (43), above, the ceratobranchial (42), forming the middle part, and the hypobranchial (41), the lower part articulating with the series of basibranchials (40) which lie behind the epihyal of the tongue. On the three bones forming the first gill-arch are attached numerous appendages called gill-rakers (47). These gill-rakers vary very greatly in number and form. In the striped bass they are few and spear-shaped. In the shad they are very many and almost as fine as hairs. In some fishes they form an effective strainer in separating the food, or perhaps in keeping extraneous matter from the gills. In some fishes they are short and lumpy, in others wanting altogether.
The anterior vertebræ known as abdominal vertebræ, bounding the body-cavity, possess neural spines similar to those of the caudal vertebræ. In place, however, of the hæmapophyses are projections known as parapophyses (72), which do not meet below, but extend outward, forming the upper part of the wall of the abdominal cavity.
In the garpike the vertebræ are convex anteriorly, concave behind, being joined by ball-and-socket joints (opisthocœlian). In most other fishes they are double concave (amplicœlian). In sharks the vertebræ are imperfectly ossified, a number of terms, asterospondylous, cyclospondylous, tectospondylous, being applied to the different stages of ossification, these terms referring to the different modes of arrangement of the calcareous material within the vertebra.
The post-temporal sometimes projects behind through the skin and may bear spines or serrations. In front of the post-temporal and a little to the outside of it is the small supratemporal (52) also usually connecting the shoulder-girdle with the skull. Below the post-temporal, extending downward and backward, is the flattish supraclavicle (posterotemporal) (54). To this is joined the long clavicle (proscapula) (55), which runs forward and downward in the bony fishes, meeting its fellow on the opposite side in a manner suggesting the wishbone of a fowl. Behind the base of the clavicle, the sword-shaped post-clavicle (56) extends downward through the muscles behind the base of the pectoral fin. In some fishes, as the stickleback and the trumpet-fish, a pair of flattish or elongate bones called interclavicles (infraclavicles) lie between and behind the lower part of the clavicle. These are not found in most fishes and are wanting in the striped bass. They are probably in all cases merely extensions of the hypocoracoid.
mesocoracoid
In the shark the pelvic girdle is rather largely developed, but in the more specialized fishes it loses its importance. In the less specialized of the bony fishes the pelvis is attached at a distance from the head among the muscles of the side, and free from the shoulder-girdle and other parts of the skeleton. The ventral fins are then said to be abdominal. When very close to the clavicle, but not connected with it, as in the mullet, the fin is still said to be abdominal or subabdominal. In the striped bass the pelvis is joined by ligament between the clavicles, near their tip. The ventral fins thus connected, as seen in most spiny-rayed fishes, are said to be thoracic. In certain forms the pelvis is thrown still farther forward and attached at the throat or even to the chin. When the ventral fins are thus inserted before the shoulder-girdle, they are said to be jugular. Most of the fishes with spines in the fins have thoracic ventrals. In the fishes with jugular ventrals these fins have begun a process of degeneration by which the spines or soft rays or both are lost or atrophied.
The Skeleton in Primitive Fishes.—To learn the names of bones we can deal most satisfactorily with the higher fishes, those in which the bony framework has attained completion. But to understand the origin and relation of parts we must begin with the lowest types, tracing the different stages in the development of each part of the system.
In the embryo of the bony fish a similar notochord precedes the segmentation and ossification of the vertebral column. In most of the extinct types of fishes a notochord more or less modified persisted through life, the vertebræ being strung upon it spool fashion in various stages of development. In the Cyclostomi (lampreys and hagfishes) the limbs and lower jaw are still wanting, but a distinct skull is developed. The notochord is still present, but its anterior pointed end is wedged into the base of a cranial capsule, partly membranous, partly cartilaginous. There is no trace of segmentation in the notochord itself in these or any other fishes, but neutral arches are foreshadowed in a series of cartilages on each side of the spinal chord. The top of the head is protected by broad plates. There are ring-like cartilages supporting the mouth and other cartilages in connection with the tongue and gill structures.
The shoulder-girdle is made of a single cartilage, touching the back-bone at a distance behind the head. To this cartilage three smaller ones are attached, forming the base of the pectoral fin. These are called mesopterygium, propterygium, and metapterygium, the first named being in the middle and more distinctly basal. These three segments are subject to much variation. Sometimes one of them is wanting; sometimes two are grown together. Behind these the fin-rays are attached. In most of the skates the shoulder-girdle is more closely connected with the anterior vertebræ, which are more or less fused together.
Evidence of Paleontology.—If we had representations of all the early forms of fishes arranged in proper sequence, we could decide once for all, by evidence of paleontology, which form of fin appears first and what is the order of appearance. As to this, it is plain that we do not know the most primitive form of fin. Sharks of unknown character must have existed long before the earliest remains accessible to us. Hence the evidence of paleontology seems conflicting and uncertain. On the whole it lends most support to the fin-fold theory. In the later Devonian, a shark, Cladoselache fyleri, is found in which the paired fins are lappet-shaped, so formed and placed as to suggest their origin from a continuous fold of skin. In this species the dorsal fins show much the same form. Other early sharks, constituting the order of Acanthodei, have fins somewhat similar, but each preceded by a stiff spine, which may be formed from coalescent rays.
Long after these appears another type of sharks represented by Pleuracanthus and Cladodus, in which the pectoral fin is a jointed organ fringed with rays arranged serially in one or two rows. This form of fin has no resemblance to a fold of skin, but accords better with Gegenbaur's theory that the pectoral limb was at first a modified gill-arch. In the Coal Measures are found also teeth of sharks (Orodontidæ) which bear a strong resemblance to still existing forms of the family of Heterodontidæ, which originates in the Permian. The existing Heterodontidæ have the usual specialized form of shark-fin, with three of the basal segments especially enlarged and placed side by side, the type seen in modern sharks. Whatever the primitive form of shark-fin, it may well be doubted whether any one of these three (Cladoselache, Pleuracanthus, or Heterodontus) actually represents it. The beginning is therefore unknown, though there is some evidence that Cladoselache is actually more nearly primitive than any of the others. As we shall see, the evidence of comparative anatomy may be consistent with either of the two chief theories, while that of ontogeny or embryology is apparently inconclusive, and that of paleontology is apparently most easily reconciled with the theory of the fin-fold.
Current Theories as to Origin of Paired Fins.—There are three chief theories as to the morphology and origin of the paired fins. The earliest is that of Dr. Karl Gegenbaur, supported by various workers among his students and colleagues. In his view the pectoral and ventral fins are derived from modifications of primitive gill-arches. According to this theory, the skeletal arrangements of the vertebrate limb are derived from modifications of one primitive form, a structure made up of successive joints, with a series of fin-rays on one or both sides of it. To this structure Gegenbaur gives the name of archipterygium. It is found in the shark, Pleuracanthus, in Cladodus, and in all the Dipnoan and Crossopterygian fishes, its primitive form being still retained in the Australian genus of Dipnoans, Neoceratodus. This biserial archipterygium with its limb-girdle is derived from a series of gill-rays attached to a branchial arch. The backward position of the ventral fin is due to a succession of migrations in the individual and in the species.
"This view is based on the skeletal structures within the fin. It rests upon (1) the assumption that the archipterygium is the primitive type of fin, and (2) the fact that amongst the Selachians is found a tendency for one branchial ray to become larger than the others, and, when this has happened, for the base of attachment of neighboring rays to show a tendency to migrate from the branchial arch on to the base of the larger or, as we may call it, primary ray; a condition coming about which, were the process to continue rather farther than it is known to do in actual fact, would obviously result in a structure practically identical with the archipterygium. Gegenbaur suggests that the archipterygium actually has arisen in this way in phylogeny."
"This view, in its modern form, was based by Balfour on his observation that in the embryos of certain Elasmobranchs the rudiments of the pectoral and pelvic fins are at a very early period connected together by a longitudinal ridge of thickened epiblast—of which indeed they are but exaggerations. In Balfour's own words referring to these observations: 'If the account just given of the development of the limb is an accurate record of what really takes place, it is not possible to deny that some light is thrown by it upon the first origin of the vertebrate limbs. The facts can only bear one interpretation, viz., that the limbs are the remnants of continuous lateral fins.'
We may now take up the evidence in regard to each of the different theories, using in part the language of Kerr, the paragraphs in quotation-marks being taken from his paper. We may first consider Balfour's theory of the lateral fold.
"The portion of Gegenbaur's view which asserts that the biserial archipterygial fin is of an extremely primitive character is supported by a large body of anatomical facts, and is rendered further probable by the great frequency with which fins apparently of this character occur amongst the oldest known fishes. On the lateral-fold view we should have to regard these as independently evolved, which would imply that fins of this type are of a very perfect character, and in that case we may be indeed surprised at their so complete disappearance in the more highly developed forms, which followed later on."
As to Gegenbaur's theory it is urged that no form is known in which a gill-septum develops into a limb during the growth of the individual. The main thesis, according to Professor Kerr, "that the archipterygium was derived from gill-rays, is supported only by evidence of an indirect character. Gegenbaur in his very first suggestion of his theory pointed out, as a great difficulty in the way of its acceptance, the position of the limbs, especially of the pelvic limbs, in a position far removed from that of the branchial arches. This difficulty has been entirely removed by the brilliant work of Gegenbaur's followers, who have shown from the facts of comparative anatomy and embryology that the limbs, and the hind limbs especially, actually have undergone, and in ontogeny do undergo, an extensive backward migration. In some cases Braus has been able to find traces of this migration as far forward as a point just behind the branchial arches. Now, when we consider the numbers, the enthusiasm, and the ability of Gegenbaur's disciples, we cannot help being struck by the fact that the only evidence in favor of this derivation of the limbs has been that which tends to show that a migration of the limbs backwards has taken place from a region somewhere near the last branchial arch, and that they have failed utterly to discover any intermediate steps between gill-rays and archipterygial fin. And if for a moment we apply the test of common sense we cannot but be impressed by the improbability of the evolution of a gill-septum, which in all the lower forms of fishes is fixed firmly in the body-wall, and beneath its surface, into an organ of locomotion.
"No doubt these external gills are absent also in a few of the admittedly primitive forms such as, e.g., (Neo-) Ceratodus. But I would ask that in this connection one should bear in mind one of the marked characteristics of external gills—their great regenerative power. This involves their being extremely liable to injury and consequently a source of danger to their possessor. Their absence, therefore, in certain cases may well have been due to natural selection. On the other hand, the presence in so many lowly forms of these organs, the general close similarity in structure that runs through them in different forms, and the exact correspondence in their position and relations to the body can, it seems to me, only be adequately explained by looking on them as being homologous structures inherited from a common ancestor and consequently of great antiquity in the vertebrate stem."
To the same type of tail Mr. Alexander Agassiz gave, in 1877, the name of leptocardial, on the supposition that it represented the adult condition of the lancelet. In this creature, however, rudimentary basal rays are present, a condition differing from that of the early embryos.
The term gephyrocercal is devised by Ryder for fishes in which the end of the vertebral axis is aborted in the adult, leaving the caudal elements to be inserted on the end of this axis, thus bridging over the interval between the vertical fins, as the name (γεφύρος, bridge; κέρκος, tail) is intended to indicate. Such a tail has been recognized in four genera only, Mola, Ranzania, Fierasfer, and Echiodon, the head-fishes and the pearl-fishes.
"1. The forms that are best comparable and that are most nearly related to each other are the Dipnoi, an order of fishes at present represented by Lepidosiren, Protopterus, and Ceratodus, and the Batrachians as represented by the Ganocephala, Salamanders, and Salamander-like animals.
"The scapula in the Urodele and other Batrachians is entirely or almost wholly excluded from the glenoid foramen, and above the coracoid. Therefore the corresponding element in Dipnoi must be the scapula.
"Neither the scapula in Batrachians nor the cartilaginous extension thereof, designated suprascapula, is dissevered from the coracoid. Therefore there is an a priori improbability against the homology with the scapula of any part having a distant and merely ligamentous connection with the humerus-bearing element. Consequently, as an element better representing the scapula exists, the element named scapula (by Owen, Günther, etc.) cannot be the homologue of the scapula of Batrachians. On the other hand, its more intimate relations with the skull and the mode of development indicate that it is rather an element originating and developed in more intimate connection with the skull. It may therefore be considered, with Parker, as a post-temporal.
"With the humerus of the Dipnoans, the element of the Polypterids (single at the base, but immediately divaricating and with its limbs bordering an intervening cartilage which supports the pectoral and its basilar ossicles) must be homologous. But it is evident that the external elements of the so-called carpus of the teleosteoid Ganoids are homologous with that element in Polypterids. Therefore those elements cannot be carpal, but must represent the humerus.
The gills, or branchiæ, are primarily folds of the skin lining the branchial cavity. In most fishes they form fleshy fringes or laminæ throughout which the capillaries are distributed. In the embryos of sharks, skates, chimæras, lung-fishes, and Crossopterygians external gills are developed, but in the more specialized forms these do not appear outside the gill-cavity. In some of the sharks, and especially the rays, a spiracle or open foramen remains behind the eye. Through this spiracle, leading from the outside into the cavity of the mouth, water is drawn downwards to pass outward over the gills. The presence of this breathing-hole permits these animals to lie on the bottom without danger of inhaling sand.
The Air-bladder.—The air-bladder, or swim-bladder, must be classed among the organs of respiration, although in the higher fishes its functions in this regard are rudimentary, and in some cases it has taken collateral functions (as a hydrostatic organ of equilibrium, or perhaps as an organ of hearing) which have no relation to its original purpose.
In front of the cerebellum lies the largest pair of lobes, each of them hollow, the optic nerves being attached to the lower surface. These are known as the optic lobes, or mesencephalon. In front of these lie the two lobes of the cerebrum, also called the hemispheres, or prosencephalon. These lobes are usually smaller than the optic lobes and solid. In some fishes they are crossed by a furrow, but are never corrugated as in the brain of the higher animals. In front of the cerebrum lie the two small olfactory lobes, which receive the large olfactory nerve from the nostrils. From its lower surface is suspended the hypophysis or pituitary gland.
In most of the bony fishes the structure of the brain does not differ materially from that seen in the perch. In the sturgeon, however, the parts are more widely separated. In the Dipnoans the cerebral hemispheres are united, while the optic lobe and cerebellum are very small. In the sharks and rays the large cerebral hemispheres are usually coalescent into one, and the olfactory nerves dilate into large ganglia below the nostrils. The optic lobes are smaller than the hemispheres and also coalescent. The cerebellum is very large, and the surface of the medulla oblongata is more or less modified or specialized. The brain of the shark is relatively more highly developed than that of the bony fishes, although in most other regards the latter are more distinctly specialized.
In the lancelet there is a single median organ supposed to be a nostril, a small depression at the front of the head, covered by ciliated membrane. In the hagfish the single median nostril pierces the roof of the mouth, and is strengthened by cartilaginous rings, like those of the windpipe. In the lamprey the single median nostril leads to a blind sac. In the Barramunda (Neoceratodus) there are both external and internal nares, the former being situated just within the upper lip. In all other fishes there is a nasal sac on either side of the head. This has usually, but not always, two openings.
The Organs of Sight.—The eyes of fishes differ from those of the higher vertebrates mainly in the spherical form of the crystalline lens. This extreme convexity is necessary because the lens itself is not very much denser than the fluid in which the fishes live. The eyes vary very much in size and somewhat in form and position. They are larger in fishes living at a moderate depth than in shore fishes or river fishes. At great depths, as a mile or more, where all light is lost, they may become aborted or rudimentary, and may be covered by the skin. Often species with very large eyes, making the most of a little light or of light from their own luminous spots, will inhabit the same depths with fishes having very small eyes or eyes apparently useless for seeing, retained as vestigial structures through heredity. Fishes which live in caves become also blind, the structures showing every possible phase of degradation. The details of this gradual loss of eyes, whether through reversed selection or hypothetically through inheritance of atrophy produced by disuse, have been given in a number of memoirs on the blind fishes of the Mississippi Valley by Dr. Carl H. Eigenmann.
In the eyes of the fishes there is no lachrymal gland. True eyelids no fishes possess; the integuments of the head pass over the eye, becoming transparent as they cross the orbit. In some fishes part of this integument is thickened, covering the eye fully although still transparent. This forms the adipose eyelid characteristic of the mullet, mackerel, and ladyfish. Many of the sharks possess a distinct nictitating membrane or special eyelid, moved by a set of muscles. The iris in most fishes surrounds a round pupil without much power of contraction. It is frequently brightly colored, red, orange, black, blue, or green. In fishes, like rays or flounders, which lie on the bottom, a dark lobe covers the upper part of the pupil—a curtain to shut out light from above. The cornea is little convex, leaving small space for aqueous humor. In two genera of fishes, Anableps, Dialommus, the cornea is divided by a horizontal partition into two parts. This arrangement permits these fishes, which swim at the surface of the water, to see both in and out of the medium. Anableps, the four-eyed fish, is a fresh-water fish of tropical America, which swims at the surface like a top-minnow, feeding on insects. Dialommus is a marine blenny from the Panama region, apparently of similar habit.
The Organs of Hearing.—The ear of the typical fish consists of the labyrinth only, including the vestibule and usually three semicircular canals, these dilating into sacs which contain one or more large, loose bones, the ear-stones or otoliths. In the lampreys there are two semicircular canals, in the hagfish but one. There is no external ear, no tympanum, and no Eustachian tube. The ear-sac on each side is lodged in the skull or at the base of the cranial cavity. It is externally surrounded by bone or cartilage, but sometimes it lies near a fontanelle or opening in the skull above. In some fishes it is brought into very close connection with the anterior end of the air-bladder. The latter organ it is thought may form part of the apparatus for hearing. The arrangement for this purpose is especially elaborate in the carp and the catfish families. In these fishes and their relatives (called Ostariophysi) the two vestibules are joined in a median sac (sinus impar) in the substance of the basioccipital. This communicates with two cavities in the atlas, which again are supported by two small bones, these resting on a larger one in connection with the front of the air-bladder. The system of bones is analogous to that found in the higher vertebrates, but it connects with the air-bladder, not with an external tympanum. The bones are not homologous with those of the ear of higher animals, being processes of the anterior vertebræ. The tympanic chain of higher vertebrates has been thought homologous with the suspensory of the mandible.
The Sense of Touch.—The sense of touch is better developed among fishes. Most of them flee from contact with actively moving objects. Many fishes use sensitive structures as a means of exploring the bottom or of feeling their way to their food. The barbel or fleshy filament wherever developed is an organ of touch. In some fishes, barbels are outgrowths from the nostrils. In the catfish the principal barbel grows from the rudimentary maxillary bone. In the horned dace and gudgeon the little barbel is attached to the maxillary. In other fishes barbels grow from the skin of the chin or snout. In the goatfish and surmullet the two chin barbels are highly specialized. In Polymixia the chin barbels are modified branchiostegals. In the codfish the single beard is little developed. In the gurnards and related forms the lower rays of the pectoral are separate and barbel-like. Detached rays of this sort are found in the thread-fins (Polynemidæ), the gurnards (Triglidæ), and in various other fishes. Barbels or fleshy flaps are often developed over the eyes and sometimes on the scales or the fins.
In others, as the surf-fishes (Embiotocidæ), they are relatively large and few in number. In the viviparous sharks, which constitute the majority of the species of living sharks, the young are large at birth and prepared to take care of themselves.
In the higher fishes the eggs are spherical, large or small according to the species, and varying in the firmness of their outer walls. All contain food-yolk from which the embryo in its earlier stages is fed. The eggs of the eel (Anguilla) are microscopic. According to Günther 25,000 eggs have been counted in the herring, 155,000 in the lumpfish, 3,500,000 in the halibut, 635,200 in the sturgeon, and 9,344,000 in the cod. Smaller numbers are found in fishes with large ova. The red salmon has about 3500 eggs, the king salmon about 5200. Where an oviduct is present the eggs are often poured out in glutinous masses, as in the bass. When, as in the salmon, there is no oviduct, the eggs lie separate and do not cohere together. It is only with the latter class of fishes, those in which the eggs remain distinct, that artificial impregnation and hatching is practicable. In this regard the value of the salmon and trout is predominant. In some fishes, especially those of elongate form, as the needle-fish (Tylosurus), the ovary of but one side is developed.
The Significance of Facts of Development.—The significance of the process of development in any species is yet far from completely understood. It is believed that many of the various stages in the development of an animal correspond to or repeat the structural condition of the animal's ancestors. Naturalists believe that all animals having a notochord at any stage in their existence are related to each other through being descended from a common ancestor, the first or oldest chordate or back-boned animal. In fact it is because all these chordate animals—the lancelets, lampreys, fishes, batrachians, the reptiles, the birds, and the mammals—have descended from a common ancestor that they all develop a notochord, and those most highly organized replace this by a complete back-bone. It is believed that the descendants of the first back-boned animal have, in the course of many generations, branched off little by little from the original type until there came to exist very real and obvious differences among the back-boned animals—differences which among the living back-boned animals are familiar to all of us. The course of development of an individual animal is believed to be a very rapid and evidently much condensed and changed recapitulation of the history which the species or kind of animal to which the developing individual belongs has passed through in the course of its descent through a long series of gradually changing ancestors. If this is true, then we can readily understand why the fish and the salamander and the tortoise and bird and rabbit are all alike in their earlier stages of development, and gradually come to differ more and more as they pass through later and later developmental stages.
Aside from the question of periblast, the growth of the blastoderm appears not unlike that of the sturgeon. From the blastula stage to that of the early gastrula, the changes have been but slight; the blastoderm has greatly flattened out as its margins grow downward, leaving the segmentation cavity apparent. The rim of the blastoderm has become thickened as the 'germ-ring'; and immediately in front of the dorsal lip of the blastopore its thickening marks the appearance of the embryo. The germ-ring continues to grow downward, and shows more prominently the outline of the embryo; this now terminates at the head region; while on either side of this point spreads out tail-ward on either side the indefinite layer of outgrowing
mesoderm. In the next stage the closure of the blastopore
is rapidly becoming completed; in front of it stretches the widened and elongated form of the embryo. The yolk-plug is next replaced by periblast, the dorsal lip by the tail-mass, or more accurately the dorsal section of the germ-rim; the cœlenteron under the dorsal lip has here disappeared, on account of the close approximation of the embryo to the periblast; its last remnant, the Kupffer's vesicle, is shortly to disappear. The germ-layers become confluent, but, unlike the sturgeon, the flattening of the dorsal germ-ring does not permit the formation of a neurenteric canal.
The mode of formation of the gill-slit of the Teleost does not differ from that in other groups; an evagination of the entoderm coming in contact with an invaginated tract of ectoderm fuses, and at this point an opening is later established.
The fish may, in general, be said to begin its existence as a larva as soon as it emerges from its egg-membranes. In some instances, however, it is difficult to decide at what point the larval stage is actually initiated: thus in sharks the excessive amount of yolk material which has been provided for the growth of the larva renders unnecessary the emerging from the egg at an early stage; and the larval period is accordingly to be traced back to stages that are still enclosed in the egg-membranes. In all cases the larval life may be said to begin when the following conditions have been fulfilled: the outward form of the larva must be well defined, separating it from the mass of yolk, its motions must be active, it must possess a continuous vertical fin-fold passing dorsally from the head region to the body terminal, and thence ventrally as far as the yolk region; and the following structures, characteristic in outward appearance, must also be established: the sense-organs—eye, ear, and nose—mouth and anus, and one or more gill-clefts.
Peculiar Larval Forms.—The young fish usually differs from the adult mainly in size and proportions. The head is larger in the young, the fins are lower, the appendages less developed, and the body more slender in the young than in the adult. But to most of these distinctions there are numerous exceptions, and in some fish there is a change so marked as to be fairly called a metamorphosis. In such cases the young fish in its first condition is properly called a larva. The larva of the lamprey (Petromyzon) is nearly blind and toothless, with slender head, and was long supposed to belong to a different genus (Ammocœtes) from the adult. The larva of sharks and rays, and also of Dipnoans and Crossopterygians, are provided with bushy external gills, which disappear in the process of development. In most soft-rayed fishes the embryonic fringe which precedes the development of the vertical fins persists for a considerable time. In many young fishes, especially the Chætodontidæ and their allies (butterfly-fishes), the young fish has the head armed with broad plates formed by the backward extension of certain membrane-bones. In other forms the bones of the head are in the young provided with long spines or with serrations, which vanish totally with age. Such a change is noticeable in the swordfish. In this species the production of the bones of the snout and upper jaw into a long bony sword, or weapon of offense, takes place only with age. The young fish have jaws more normally formed, and armed with ordinary teeth. In the headfish (Mola mola) large changes take place in the course of growth, and the young have been taken for a different type of fishes. Among certain soft-rayed fishes and eels the young is often developed in a peculiar way, being very soft, translucent, or band-like, and formed of large or loosely aggregated cells. These peculiar organisms, long known as leptocephali, have been shown to be the normal young of fishes when mature very different. In the ladyfish (Albula) Dr. Gilbert has shown, by a full series of specimens, that in their further growth these pellucid fishes shrink in size, acquiring greater compactness of body, until finally reaching about half their maximum length as larvæ. After this, acquiring essentially the form of the adult fish, they begin a process of regular growth. This leptocephalous condition is thought by Günther to be due to arrest of growth in abnormal individuals, but this is not the case in Albula, and it is probably fully normal in the conger and other eels. In the surf-fishes the larvæ have their vertical fins greatly elevated, much higher than in the adult, while the body is much more closely compressed. In the deal-fish (Trachypterus) the form of the body and fins changes greatly with age, the body becoming more elongate and the fins lower. The differences between different stages of the same fish seem greater than the differences between distinct species. In fact with this and with other forms which change with age, almost the only test of species is found in the count of the fin-rays. So far as known the numbers of these structures do not change. In the moonfishes (Carangidæ) the changes with age are often very considerable. We copy Lütken's figure of the changes in the genus Selene (fig. 113). Similar changes take place in Alectis, Vomer, and other genera.
The Age of Fishes.—The age of fishes is seldom measured by a definite period of years. Most of them grow as long as they live, and apparently live until they fall victims to some stronger species. It is reputed that carp and pike have lived for a century, but the evidence needs verification. Some fishes, as the salmon of the Pacific (Oncorhynchus), have a definite period of growth (usually four years) before spawning. After this act all the individuals die so far as known. In Japan and China the Ice-fish (Salanx), a very long, slender, transparent fish allied to the trout, may possibly be annual in habit, all the individuals perhaps dying in the fall to be reproduced from eggs in the spring. But this alleged habit needs verification.
Some fresh-water fishes are very fragile, dying soon in the air, often with injured air-bladder or blood-vessels. They will die even sooner in foul water. Other fishes are extremely tenacious of life. The mud-minnow (Umbra) is sometimes ploughed up in the half-dried mud of Wisconsin prairies. The related Alaskan blackfish (Dallia) has been fed frozen to dogs, escaping alive from their stomachs after being thawed out. Many of the catfishes (Siluridæ) will live after lying half-dried in the dust for hours. The Dipnoan, Lepidosiren, lives in a ball of half-dried mud during the arid season, and certain fishes, mostly Asiatic, belonging to the group Labyrinthici, with accessory breathing organ can long maintain themselves out of water. Among these is the China-fish (Ophiocephalus), often kept alive in the Chinese settlements in California and Hawaii. Some fishes can readily endure prolonged hunger, while others succumb as readily as a bird or a mammal.
The Effects of Temperature on Fish.—The limits of distribution of many fishes are marked by changes in temperature. Few marine fishes can endure any sudden or great change in this regard, although fresh-water fishes adapt themselves to the seasons. I have seen the cutlass-fish (Trichiurus) benumbed with cold off the coast of Florida while the temperature was still above the frost-line. Those fishes which are tenacious of life and little sensitive to changes in climate and food are most successfully acclimatized or domesticated. The Chinese carp (Cyprinus carpio) and the Japanese goldfish (Carassius auratus) have been naturalized in almost all temperate and tropical river basins. Within the limits of clear, cold waters most of the salmon and trout are readily transplanted. But some similar fishes (as the grayling) are very sensitive to the least change in conditions. Most of the catfish (Siluridæ) will thrive in almost any fresh waters except those which are very cold.
The movements of many migratory animals are mainly controlled by the impulse to reproduce. Some pelagic fishes, especially flying fishes and fishes allied to the mackerel, swim long distances to a region favorable for a deposition of spawn. Some species are known only in the waters they make their breeding homes, the individuals being scattered through the wide seas at other times. Many fresh-water fishes, as trout, suckers, etc., forsake the large streams in the spring, ascending the small brooks where they can rear their young in greater safety. Still others, known as anadromous fishes, feed and mature in the sea, but ascend the rivers as the impulse of reproduction grows strong. An account of these is given in a subsequent paragraph.
Adaptation to Environment.—In general food-securing structures are connected with the mouth, or, as in the anglers, are hung as lures above it; spines of offense and defense, electric organs, poison-glands, and the like are used in self-protection; the bright nuptial colors and adornments of the breeding season are doubtfully classed as useful in rivalry; the egg-sacs, nests, and other structures or habits may serve to defend the young, while skinny flaps, sand or weed-like markings, and many other features of mimicry serve as concessions to the environment.
The flight of one of the largest flying fishes (Cypsilurus californicus) has been carefully studied by Dr. Charles H. Gilbert and the writer. The movements of the fish in the water are extremely rapid. The sole motive power is the action under the water of the strong tail. No force can be acquired while the fish is in the air. On rising from the water the movements of the tail are continued until the whole body is out of the water. When the tail is in motion the pectorals seem in a state of rapid vibration. This is not produced by muscular action on the fins themselves. It is the body of the fish which vibrates, the pectorals projecting farthest having the greatest amplitude of movement. While the tail is in the water the ventral fins are folded. When the action of the tail ceases the pectorals and ventrals are spread out wide and held at rest. They are not used as true wings, but are held out firmly, acting as parachutes, enabling the body to skim through the air. When the fish begins to fall the tail touches the water. As soon as it is in the water it begins its motion, and the body with the pectorals again begins to vibrate. The fish may, by skimming the water, regain motion once or twice, but it finally falls into the water with a splash. While in the air it suggests a large dragon-fly. The motion is very swift, at first in a straight line, but is later deflected in a curve, the direction bearing little or no relation to that of the wind. When a vessel passes through a school of these fishes, they spring up before it, moving in all directions, as grasshoppers in a meadow.
Quiescent Fishes.—Some fishes, as the lancelet, lie buried in the sand all their lives. Others, as the sand-darter (Ammocrypta pellucida) and the hinalea (Julis gaimard), bury themselves in the sand at intervals or to escape from their enemies. Some live in the cavities of tunicates or sponges or holothurians or corals or oysters, often passing their whole lives inside the cavity of one animal. Many others hide themselves in the interstices of kelp or seaweeds. Some eels coil themselves in the crevices of rocks or coral masses, striking at their prey like snakes. Some sea-horses cling by their tails to gulfweed or sea-wrack. Many little fishes (Gobiomorus, Carangus, Psenes) cluster under the stinging tentacles of the Portuguese man-of-war or under ordinary jellyfishes. In the tide-pools, whether rock, coral, or mud, in all regions multitudes of little fishes abound. As these localities are neglected by most collectors, they have proved of late years a most prolific source of new species. The tide-pools of Cuba, Key West, Cape Flattery, Sitka, Unalaska, Monterey, San Diego, Mazatlan, Hilo, Kailua and Waiahæ in Hawaii, Apia and Pago-Pago in Samoa, the present writer has found peculiarly rich in rock-loving forms. Even richer are the pools of the promontories of Japan, Hakodate Head, Misaki, Awa, Izu, Waka, and Kagoshima, where a whole new fish fauna unknown to collectors in markets and sandy bays has been brought to light. Some of these rockfishes are left buried in the rock weeds as the tide flows, lying quietly until it returns. Others cling to the rocks by ventral suckers, while still others depend for their safety on their powers of leaping or on their quickness of their movements in the water. Those of the latter class are often brilliantly colored, but the others mimic closely the algæ or the rocks. Some fishes live in the sea only, some prefer brackish-water. Some are found only in the rivers, and a few pass more or less indiscriminately from one kind of water to another.
Anadromous Fishes.—Many fresh-water fishes, as trout and suckers, forsake the large streams in the spring, ascending the small brooks where their young can be reared in greater safety. Still others, known as anadromous fishes, feed and mature in the sea, but ascend the rivers as the impulse of reproduction grows strong. Among such fishes are the salmon, shad, alewife, sturgeon, and striped bass in American waters. The most remarkable case of the anadromous instinct is found in the king salmon or quinnat (Oncorhynchus tschawytscha) of the Pacific Coast. This great fish spawns in November, at the age of four years and an average weight of twenty-two pounds. In the Columbia River it begins running with the spring freshets in March and April. It spends the whole summer, without feeding, in the ascent of the river. By autumn the individuals have reached the mountain streams of Idaho, greatly changed in appearance, discolored, worn, and distorted. The male is humpbacked, with sunken scales, and greatly enlarged, hooked, bent, or twisted jaws, with enlarged dog-like teeth. On reaching the spawning beds, which may be a thousand miles from the sea in the Columbia, over two thousand in the Yukon, the female deposits her eggs in the gravel of some shallow brook. The male covers them and scrapes the gravel over them. The female salmon does as much as the male in covering the eggs. Then both male and female drift tail foremost helplessly down the stream; none, so far as certainly known, ever survive the reproductive act. The same habits are found in the five other species of salmon in the Pacific, but in most cases the individuals do not start so early nor run so far. The blue-back salmon or redfish, however, does not fall far short in these regards. The salmon of the Atlantic has a similar habit, but the distance traveled is everywhere much less, and most of the hook-jawed males drop down to the sea and survive to repeat the acts of reproduction.
It is well known that in the very young flounder the body rests upright in the water. After a little there is a tendency to turn to one side and the lower eye begins its migration to the other side, the interorbital bones or part of them moving before it. In most flounders the eye seems to move over the surface of the head, before the dorsal fin, or across the axil of its first ray. In the tropical genus Platophrys the movement of the eye is most easily followed, as the species reach a larger size than do most flounders before the change takes place. The larva, while symmetrical, is in all cases transparent.
4. Correlated with this is an increase in distance between the eyes and the brain, caused by the growth of the facial cartilages.
In Thalassophryne, a genus of toad-fishes of tropical America, is found the most perfect system of poison organs known among fishes. The spinous armature of the opercle and the two spines of the first dorsal fin constitute the weapons. The details are known from the dissections of Dr. Günther. According to his[19] observations, the opercle in Thalassophryne "is very narrow, vertically styliform and very mobile. It is armed behind with a spine eight lines long and of the same form as the hollow venom-fang of a snake, being perforated at its base and at its extremity. A sac covering the base of the spine discharges its contents through the apertures and the canal in the interior of the spine. The structure of the dorsal spines is similar. There are no secretory glands imbedded in the membranes of the sacs and the fluid must be secreted by their mucous membrane. The sacs are without an external muscular layer and situated immediately below the thick, loose skin which envelops the spines at their extremity. The ejection of the poison into a living animal, therefore, can only be effected as in Synanceia, by the pressure to which the sac is subjected the moment the spine enters another body."
The flesh of the filefishes (Stephanolepis tomentosus), which the writer has tested, is very meager and bitter, having a decidedly offensive taste. It is suspected, probably justly, of being poisonous. In the globefishes the flesh is always more or less poisonous, that of Tetraodon hispidus, called muki-muki, or death-fish, in Hawaii, is reputed as excessively so. The poisonous fishes have been lately studied in detail by Dr. Jacques Pellegrin, of the Museum d'Histoire Naturelle at Paris. He shows that any species of fish may be poisonous under certain circumstances, that under certain conditions certain species are poisonous, and that certain kinds are poisonous more or less at all times. The following account is condensed from Dr. Pellegrin's observations.
It is claimed in tropical regions that the germs of the bubonic plague may be carried through the mediation of fishes which feed on sewage. It is suggested by Dr. Charles B. Ashmead that leprosy may be so carried. It is further suggested that the custom of eating the flesh of fishes raw almost universal in Japan, Hawaii, and other regions may be responsible for the spread of certain contagious diseases, in which the fish acts as an intermediate host, much as certain mosquitoes spread the germ of malaria and yellow fever.
The electric eel (Electrophorus electricus), the most powerful of electric fishes, is not an eel, but allied rather to the sucker or carp. It is, however, eel-like in form and lives in rivers of Brazil and Guiana. The electric organs are in two pairs, one on the back of the tail, the other on the anal fin. These are made up of an enormous number of minute cells. In the electric eel, as in the other electric fishes, the nerves supplying these organs are much larger than those passing from the spinal cord for any other purpose. In all these cases closely related species show a no trace of the electric powers.
Photophores or Luminous Organs.—Many fishes, chiefly of the deep seas, develop organs for producing light. These are known as luminous organs, phosphorescent organs, or photophores. These are independently developed in four entirely unrelated groups of fishes. This difference in origin is accompanied by corresponding difference in structure. The best-known type is found in the Iniomi, including the lantern-fishes and their many relatives. These may have luminous spots, differentiated areas round or oblong which shine star-like in the dark. These are usually symmetrically placed on the sides of the body. They may have also luminous glands or diffuse areas which are luminous, but which do not show the specialized structure of the phosphorescent spots. These glands of similar nature to the spots are mostly on the head or tail. In one genus, Æthoprora, the luminous snout is compared to the headlight of an engine.
In all these structures, according to Dr. von Lendenfeld, the whole or part of the organ is glandular. The glandular part is at the base and the other structures are added distally. The primitive organ was a gland which produced luminous slime. To this in the process of specialization greater complexity has been added.
"Electrical stimulation of the live fish was also tried with good success. The interrupted current from an induction coil was used, one electrode being fixed on the head over the brain or on the exposed spinal cord near the brain, and the other moved around on different parts of the body. No results followed relatively weak stimulation of the fish, although such currents produced violent contractions of the muscular system of the body. But when a current strong enough to be quite painful to the hands while handling the electrodes was used then stimulation of the fish called forth a brilliant glow of light apparently from every well-developed photophore. All the lines on the ventral and lateral surfaces of the body glowed with a beautiful light, and continued to do so while the stimulation lasted. The single well-developed organ just back of and below the eye was especially prominent. No luminosity was observed in the region of the dorsal organs previously described as rudimentary in structure. I was also able to produce the same effect by galvanic stimulation, rapidly making and breaking the current by hand.
"An important fact brought out by the above experiment is that an electrical stimulation strong enough to most violently stimulate the nervous system, as shown by the violent contractions of the muscular system, may still be too weak to produce phosphorescence. This fact gives a physiological confirmation of the morphological result stated above that no specific nerves are distributed to the phosphorescent organs.
"The experiments just related were all tried on specimens of the fish taken from under the rocks where they were guarding the young brood. Two specimens, however, taken by hooks from the deeper water of Monterey Bay, could not be made to show phosphorescence either by electrical stimulation or by treatment with ammonia. These specimens did net have the high development of the system of mucous cells of the skin exhibited by the nesting fish. My observations were, however, not numerous enough to more than suggest the possibility of a seasonal high development of the phosphorescent organs.
"In the phosphorescent organs of Porichthys the only part the function of which cannot be explained on physical grounds is the group of cells called the gland. If the large granular cells of this portion of the structure produce a secretion, as seems probable from the character of the cells and their behavior toward reagents, and this substance be oxidizable and luminous in the presence of free oxygen, i.e., photogenic, then we have the conditions necessary for a light-producing organ. The numerous capillaries distributed to the gland will supply free oxygen sufficient to meet the needs of the case. Light produced in the gland is ultimately all projected to the exterior, either directly from the luminous points in the gland or reflected outward by the reflector, the lens condensing all the rays into a definite pencil or slightly diverging cone. This explanation of the light-producing process rests on the assumption of a secretion product with certain specific characters. But comparing the organ with structures known to produce such a substance, i.e., the glands of the firefly or the photospheres of Euphausia, it seems to me the assumption is not less certain than the assumption that twelve structures resembling each other in certain particulars have a common function to that proved for one only of the twelve.
Remoras.—The different species of Remora, or shark-suckers, fasten themselves to the surface of sharks or other fishes and are carried about by them often to great distances. These fishes attach themselves by a large sucking-disk on the top of the head, which is a modified spinous dorsal fin. They do not harm the shark, except possibly to retard its motion. If the shark is caught and drawn out of the water, these fishes often instantly let go and plunge into the sea, swimming away with great celerity.
The hagfishes (Myxine, Eptatretus) alone among fishes are truly parasitic. These fishes, worm-like in form, have round mouths, armed with strong hooked teeth. They fasten themselves at the throats of large fishes, work their way into the muscle without tearing the skin, and finally once inside devour all the muscles of the fish, leaving the skin unbroken and the viscera undisturbed. These fishes become living hulks before they die. If lifted out of the water, the slimy hagfish at once slips out and swims quickly away. In gill-nets in Monterey Bay great mischief is done by hagfish (Polistotrema stouti). It is a curious fact that large numbers of hagfish eggs are taken from the stomachs of the male hagfish, which seems to be almost the only enemy of his own species, keeping the numbers in check.
It has been known for some years that in several groups of fishes (wrasse-fishes, flounders, and "rock-cod," for example) those species which inhabit northern waters have more vertebræ than those living in the tropics. Certain arctic flounders, for example, have sixty vertebræ; tropical flounders have, on the average, thirty. The significance of this fact is the problem at issue. In science it is assumed that all facts have significance, else they would not exist. It becomes necessary, then, to find out first just what the facts are in this regard.
It is certain that the possession of fewer vertebræ indicates the higher rank, the greater specialization of parts, even though the many vertebræ be a feature less primitive. The evolution of fishes is rarely a movement of progress toward complexity. The time movement in some groups is accompanied by degradation and loss of parts, by vegetative repetition of structures, and often by a movement from the fish-form toward the eel-form. Water life is less exacting than land life, having less variation of conditions. It is, therefore, less effective in pushing forward the differentiation of parts. When vertebræ are few in number each one is relatively larger, its structure is more complicated, its appendages larger and more useful, and the fins with which it is connected are better developed. In other words, the tropical fish is more intensely and compactly a fish, with a better fish equipment, and in all ways better fitted for the business of a fish, especially for that of a fish that stays at home.
The conditions most favorable to fish life are among the rocks and reefs of the tropical seas. About the coral reefs is the center of fish competition. A coral archipelago is the Paris of fishes. In such regions is found the greatest variety of surroundings, and therefore the greatest number of possible adjustments. The struggle is between fish and fish, not between fishes and hard conditions of life. No form is excluded from the competition. Cold, darkness, and foul water do not shut out competitors, nor does any evil influence sap the strength. The heat of the tropics does not make the sea-water hot. It is never sultry or laden with malaria.
Degeneration of Structures.—In the lancelet, which is a primitively simple organism, the various structures of the body are formed of simple tissues and in a very simple fashion. It is probable from the structure of each of these that it has never been very much more complex. As the individual develops in the process of growth each organ goes as it were straight to its final form and structure without metamorphosis or especial alterations by the way. When this type of development occurs, the organism belongs to a type which is primitively simple. But there are other forms which in their adult state appear feeble or simple, in which are found elements of organs of high complexity. Thus in the sea-snail (Liparis), small, weak, with feeble fins and flabby skin, we find the essential anatomy of the sculpin or the rosefish. The organs of the latter are there, but each one is reduced or degenerate, the bones as soft as membranes, the spines obsolete or buried in the skin. Such a type is said to be degenerate. It is very different from one primitively simple, and it is likely in its earlier stages of development to be more complex than when it is fully grown.
In the evolution of groups of fishes it is a common feature that some one organ will be the center of a special stress, in view of some temporary importance of its function. By the process of natural selection it will become highly developed and highly specialized. Some later changes in conditions will render this specialization useless or even harmful for at least a part of the species possessing it. The structure then undergoes degeneration, and in many cases it is brought to a lower estate than before the original changes. An example of this may be taken from the loricate or mailed-cheek fishes. One of the primitive members of this group is the rockfish known as priestfish (Sebastodes mystinus). In this fish the head is weakly armed, covered with ordinary scales. A slight suggestion of cranial ridges and a slight prolongation of the third suborbital constitute the chief suggestions of its close affinity with the mailed-cheek fishes. In other rockfishes the cranial ridges grow higher and sharper. The third suborbital extends itself farther and wider. It becomes itself spinous in still others. Finally it covers the whole cheek in a coat of mail. The head above becomes rough and horny and at last the whole body also is enclosed in a bony box. But while this specialization reaches an extraordinary degree in forms like Agonus and Peristedion, it begins to abate with Cottus, and thence through Cottunculus, Psychrolutes, Liparis, and the like, and the mailed cheek finds its final degradation in Parliparis. In this type no spines are present anywhere, no hard bone, no trace of scales, of first dorsal, or of ventral fins, and in the soft, limp structure covered with a fragile, scarf-like skin we find little suggestion of affinity with the strong rockfish or the rough-mailed Agonus. Yet a study of the skeleton shows that all these loricate forms constitute a continuous divergent series. The forms figured constitute only a few of the stages of specialization and degradation which the members of this group represent.
"(c) The third, from 10 mm. to about 80 or 100 mm. It is characterized by a number of changes which are positive as contrasted with degenerative. There are also distinct degenerative processes taking place during this period.
Equally closely will the markings on a fish correspond with rock surroundings. With granite rocks we find an elaborate series of granitic markings, with coral rocks another series of shades, and if red corals be present, red shades of like appearance are found on the fish. Still another kind of mark indicates rock pools lined with the red calcareous algæ called corallina. Black species are found in lava masses, grass-green ones among the fronds of ulva, and olive-green among Sargassum or fucus, the markings and often the form corresponding to the nature of the algæ in which the species makes its home.
Nuptial Coloration.—Nuptial colors are those which appear on the male in the breeding season only, the pigment afterwards vanishing, leaving the sexes essentially alike. Such colors are found on most of the minnows and dace (Cyprinidæ) of the rivers and to a less degree in some other fresh-water fishes, as the darters (Etheostominæ) and the trout. In the minnows of many species the male in spring has the skin charged with bright pigment, red, black, or bright silvery, for the most part, the black most often on the head, the red on the head and body, and the silvery on the tips of the fins. At the same time other markings are intensified, and in many species the head and sometimes the body and fins are covered with warty excrescences. These shades are most distinct on the most vigorous males, and disappear with the warty excrescences after the fertilization of the eggs.
Intensity of Coloration.—In general, coloration is most intense and varied in certain families of the tropical shores, and especially about coral reefs. But in brilliancy of individual markings some fresh-water fishes are scarcely less notable, especially the darters (Etheostominæ) and sunfishes (Centrarchidæ) of the streams of eastern North America. The bright hues of these fresh-water fishes are, however, more or less concealed in the water by the olivaceous markings and dark blotches of the upper parts.
Again, that which is a barrier to rapid distribution may become an agent in the slow extension of the range of a species. The great continent of Asia is undoubtedly one of the greatest of barriers to the wide movement of species of fish, yet its long shore-line enables species to creep, as it were, from bay to bay, or from rock to rock, till, in many cases, the same species is found in the Red Sea and in the tide-pools or sand-reaches of Japan. In the North Pacific, the presence of a range of half-submerged volcanoes, known as the Aleutian and the Kurile Islands, has greatly aided the slow movement of the fishes of the tide-pools and the kelp. To a school of mackerel or of flying-fishes these rough islands with their narrow channels might form an insuperable barrier.
Centers of Distribution.—We may assume, in regard to any species, that it has had its origin in or near that region in which it is most abundant and characteristic. Such an assumption must involve a very large percentage of error or of doubt, but in considering the mass of species, it may represent essential truth. In the same fashion we may regard a genus as being autochthonous or first developed in the region where it shows the greatest range or variety of species. Those regions where the greatest number of genera are thus autochthonous may be regarded as centers of distribution. So far as the marine fishes are concerned, the most important of these supposed centers are found in the Pacific Ocean. First of these in importance is the East-Indian Archipelago, with the neighboring shores of India. Next would come the Arctic Pacific and its bounding islands, from Japan to British Columbia. Third in importance in this regard is Australia. Important centers are found in temperate Japan, in California, the Panama region, and in New Zealand, Chili, and Patagonia. The fauna of Polynesia is almost entirely derived from the Indies; and the shore fauna of the Red Sea, the Bay of Bengal, and Madagascar, so far as genera are concerned, seems to be not really separable from the Indian fauna generally.
Bassalian Fishes.—The vast group of bassalian or deep-sea fishes includes those forms which live below the line of adequate light. These too are localized in their distribution, and to a much greater extent than was formerly supposed. Yet as they dwell below the influence of the sun's rays, zones and surface temperatures are nearly alike to them, and the same forms may be found in the Arctic or under the equator. Their differences in distribution are largely vertical, some living at greater depths than others, and they shade off by degrees from bathybial into semi-bathybial, and finally into ordinary pelagic and ordinary shore types. Apparently all of the bassalian fishes are derived from littoral types, the changes in structure being due to degeneration of the osseous and muscular systems and of structures not needed in deep-sea life.
Still another line of division is made by the Ural Mountains in the Old World and by the Rocky Mountains in the New. In both cases the Eastern region is vastly richer in genera and species, as well as in autochthonous forms, than the Western. The reason for this lies in the vastly greater extent of the river basins of China and the Eastern United States, as compared with those of Europe or the Californian region.
"In the appendix to the same volume Mr. Warren Upham, in discussing the probable causes of glaciation, says: 'The quaternary uplifts of the Andes and Rocky Mountains and of the West Indies make it nearly certain that the Isthmus of Panama has been similarly elevated during the recent epoch.... It may be true, therefore, that the submergence of this isthmus was one of the causes of the Glacial period, the continuation of the equatorial oceanic currents westward into the Pacific having greatly diminished or wholly diverted the Gulf Stream, which carries warmth from the tropics to the northern Atlantic and northwestern Europe.'
"If we could lower the isthmian region 300 feet at present, the waters of the two oceans would certainly commingle through the narrow Culebra Pass. But the Culebra Pass is clearly the headwater col of two streams, the Obispo flowing into the Chagres, and the Rio Grande flowing into the Pacific, and has been cut by fluviatile action, and not by marine erosion, out of a land mass which has existed since Miocene time. Those who attempt to establish Pleistocene interoceanic channels through this pass on account of its present low altitude must not omit from their calculations the restoration of former rock masses which have been removed by the general levelling of the surface by erosion."
"Prior to the elevation of a given mountain chain, the courses of the rivers may generally have been entirely different from their later ones. Subsequent to this period, they can only have undergone partial modifications. As subsequent submergences can rarely have extended to the highlands where such streams originate, the fishes of such rivers can only have been destroyed so far as they were unable to reach those elevated regions, and preserve themselves from destruction from salt water by sheltering themselves in mountain streams. On the other hand, a period of greater elevation of the land, and of consequent greater cold, would congeal the waters and cover their courses with glaciers. The fishes would be driven to the neighborhood of the coast, though no doubt in more southern latitudes a sufficient extent of uncongealed fresh waters would flow by a short course into the ocean, to preserve from destruction many forms of fresh-water fishes. Thus, through many vicissitudes, the fauna of a given system of rivers has had opportunity of uninterrupted descent, from the time of the elevation of the mountain range, in which it has its sources....
"Facts of distribution in the eastern district of North America are these. Several species of fresh-water fishes occur at the same time in many Atlantic basins from the Merrimac or from the Hudson to the James, and throughout the Mississippi Valley, and in the tributaries of the Great Lakes. On the other hand, the species of each river may be regarded as pertaining to four classes, whose distribution has direct reference to the character of the water and the food it offers: first, those of the tide-waters, of the river channels, bayous, and sluggish waters near them, or in the flat lands near the coast; second, those of the river channels of its upper course, where the currents are more distinct; third, those of the creeks of the hill country; fourth, those of the elevated mountain streams which are subject to falls and rapids."
When two or more species of any genus now inhabit the same waters, they are usually species whose differentiation is of long standing,—species, therefore, which can be readily distinguished from one another. When, on the other hand, we have "representative species,"—closely related forms, neither of which is found within the geographical range of the other,—we can with some confidence look for intermediate forms where the territory occupied by the one bounds that inhabited by the other. In very many such cases the intermediate forms have been found; and such forms are considered as subspecies of one species, the one being regarded as the parent stock, the other as an offshoot due to the influences of different environment. Then, besides these "species" and "subspecies," groups more or less readily recognizable, there are varieties and variations of every grade, often too ill-defined to receive any sort of name, but still not without significance to the student of the origin of species. Comparing a dozen fresh specimens of almost any kind of fish from any body of water with an equal number from somewhere else, one will rarely fail to find some sort of differences,—in size, in form, in color. These differences are obviously the reflex of differences in the environment, and the collector of fishes seldom fails to recognize them as such; often it is not difficult to refer the effect to the conditions. Thus fishes from grassy bottoms are darker than those taken from over sand, and those from a bottom of muck are darker still, the shade of color being, in some way not well understood, dependent on the color of the surroundings. Fishes in large bodies of water reach a larger size than the same species in smaller streams or ponds. Fishes from foul or sediment-laden waters are paler in color and slenderer in form than those from waters which are clear and pure. Again, it is often true that specimens from northern waters are less slender in body than those from farther south; and so on. Other things being equal, the more remote the localities from each other, the greater are these differences.
Local Barriers.—Least important to these are the barriers which may exist within the limits of any single basin, and which tend to prevent a free diffusion through its waters of species inhabiting any portion of it. In streams flowing southward, or across different parallels of latitude, the difference in climate becomes a matter of importance. The distribution of species is governed very largely by the temperature of the water. Each species has its range in this respect,—the free-swimming fishes, notably the trout, being most affected by it; the mud-loving or bottom fishes, like the catfishes, least. The latter can reach the cool bottoms in hot weather, or the warm bottoms in cold weather, thus keeping their own temperature more even than that of the surface of the water. Although water communication is perfectly free for most of the length of the Mississippi, there is a material difference between the faunæ of the stream in Minnesota and in Louisiana. This difference is caused chiefly by the difference in temperature occupying the difference in latitude. That a similar difference in longitude, with free water communication, has no appreciable importance, is shown by the almost absolute identity of the fish faunæ of Lake Winnebago and Lake Champlain. While many large fishes range freely up and down the Mississippi, a majority of the species do not do so, and the fauna of the upper Mississippi has more in common with that of the tributaries of Lake Michigan than it has with that of the Red River or the Arkansas. The influence of climate is again shown in the paucity of the fauna of the cold waters of Lake Superior, as compared with that of Lake Michigan. The majority of our species cannot endure the cold. In general, therefore, cold or Northern waters contain fewer species than Southern waters do, though the number of individuals of any one kind may be greater. This is shown in all waters, fresh or salt. The fisheries of the Northern seas are more extensive than those of the tropics. There are more fishes there, but are far less varied in kind. The writer once caught seventy-five species of fishes in a single haul of the seine at Key West, while on Cape Cod he obtained with the same net but forty-five species in the course of a week's work. Thus it comes that the angler, contented with many fishes of few kinds, goes to Northern streams to fish, while the naturalist goes to the South.
"Pacific Creek is a stream of good size long before it enters the pass, and its course through the meadow is in a definite channel, but not so with Atlantic Creek. The west bank of each fork is low and the stream is liable to break through anywhere and thus send part of its water across to Pacific Creek. It is probably true that one or two branches always connect the two creeks under ordinary conditions, and that following heavy rains or when the snows are melting, a much greater portion of the water of Atlantic Creek crosses the meadow to the other side.
Swampy Watersheds.—The low and irregular watershed which separates the tributaries of Lake Michigan and Lake Erie from those of the Ohio is of little importance in determining the range of species. Many of the distinctively Northern fishes are found in the headwaters of the Wabash and the Scioto. The considerable difference in the general fauna of the Ohio Valley as compared with that of the streams of Michigan is due to the higher temperature of the former region, rather than to any existing barriers between the river and the Great Lakes. In northern Indiana the watershed is often swampy, and in many places large ponds exist in the early spring.
Very near the first among sea-fishes must come the pampano (Trachinotus carolinus) of the Gulf of Mexico, with firm, white, finely flavored flesh.
Side by side with these belongs the whitefish of the Great Lakes (Coregonus clupeiformis). Its flesh, delicate, slightly gelatinous, moderately oily, is extremely agreeable. Sir John Richardson records the fact that one can eat the flesh of this fish longer than any other without the feeling of cloying. The salmon cannot be placed in the front ranks because, however excellent, the stomach soon becomes tired of it. The Spanish mackerel (Scomberomorus maculatus), with flesh at once rich and delicate, the great opah (Lampris luna), still richer and still more delicate, the bluefish (Pomatomus saltatrix) similar but a little coarser, the ulua (Carangus sem), the finest large food-fish of the South Seas, the dainty California poppy-fish, miscalled "Pampano" (Palometa simillima), and the kingfish firm and well-flavored (Scomberomorus cavalla), represent the best of the fishes allied to the mackerel.
The shad (Alosa sapidissima), with its sweet, tender, finely oily flesh, stands also near the front among food-fishes, but it sins above all others in the matter of small bones. The weak-fish (Cynoscion nobilis) and numerous relatives rank first among those with tender, white, savorous flesh. Among the bass and perch-like fishes, common consent places near the first the striped bass (Roccus lineatus), the bass of Europe (Dicentrarchus labrax), the susuki of Japan (Lateolabrax japonicus), the red tai of Japan (Pagrus major and P. cardinalis), the sheep's-head (Archosargus probatocephalus), the mutton-fish or Pargo Criollo of Cuba (Lutianus analis), the European porgy (Pagrus pagrus), the robalo (Centropomus undecimalis), the uku (Aprion virescens) of Hawaii, the spadefish (Chætodipterus faber), and the black bass (Micropterus dolomieu).
The various kinds of trout have been made famous the world over. All are attractive in form and color; all are gamey; all have the most charming of scenic surroundings, and, finally, all are excellent as food, not in the first rank perhaps, but well above the second. Notable among these are the European charr (Salvelinus alpinus), the American speckled trout or charr (Salvelinus fontinalis), the Dolly Varden or malma (Salvelinus malma), and the oquassa trout (Salvelinus oquassa). Scarcely less attractive are the true trout, the brown trout, or forelle (Salmo fario), in Europe, the rainbow-trout (Salmo irideus), the steelhead (Salmo gairdneri), the cut-throat trout (Salmo clarkii), and the Tahoe trout (Salmo henshawi), in America, and the yamabe (Salmo perryi) of Japan. Not least of all these is the flower of fishes, the grayling (Thymallus), of different species in different parts of the world.
Other most excellent food-fishes are the eel (Anguilla species), the pike (Esox lucius), the muskallonge (Esox Roccus), the sole of Europe (Solea solea), the sardine (Sardinella pilchardus), the atka-fish (Pleurogrammus monopterygius) of Bering Sea, the pescado blanco of Lake Chapala (Chirostoma estor and other species), the Hawaiian mullet (Mugil cephalus), the channel catfish (Ictalurus punctatus), the turbot (Scophthalmus maximus), the barracuda (Sphyræna), and the young of various sardines and herring, known as whitebait. Of large fishes, probably the swordfish (Xiphias gladius), the halibut (Hippoglossus hippoglossus), and the king-salmon, or quinnat (Oncorhynchus tschawytscha), may be placed first. Those people who feed on raw fish prefer in general the large parrot-fishes (as Pseudoscarus jordani in Hawaii), or else the young of mullet and similar species.
Abundance of Food-fishes.—In general, the economical value of any species depends not on its toothsomeness, but on its abundance and the ease with which it may be caught and preserved. It is said that more individuals of the herring (Clupea harengus in the Atlantic, Clupea pallasi in the Pacific) exist than of any other species. The herring is a good food-fish and whenever it runs it is freely sought. According to Björnsön, wherever the school of herring touches the coast of Norway, there a village springs up, and this is true in Scotland, Newfoundland, and from Killisnoo in Alaska to Otaru in Japan, and to Strielok in Siberia. Goode estimates the herring product of the North Atlantic at 1,500,000,000 pounds annually. In 1881 Professor Huxley used these words:
"It is said that 2,500,000,000 or thereabout of herrings are every year taken out of the North Sea and the Atlantic. Suppose we assume the number to be 3,000,000,000 so as to be quite safe. It is a large number undoubtedly, but what does it come to? Not more than that of the herrings which may be contained in one shoal, if it covers half a dozen square miles, and shoals of much larger size are on record. It is safe to say that scattered through the North Sea and the Atlantic, at one and the same time, there must be scores of shoals, any one of which would go a long way toward supplying the whole of man's consumption of herrings."
Fisheries.—With the multiplicity of apparatus for fishing, there is the greatest variety in the boats which may be used. The fishing-fleet of any port of the world is a most interesting object, as are also the fishermen with their quaint garb, plain speech, and their strange songs and calls with the hauling in of the net.
"And neither did Izaak Walton for that matter. He who is sponsor for all that is gentle in angling missed much that is best in the sport by living too early. He did not experience the exquisite pleasure of wading down mountain streams in supposedly water-proof boots and feeling the water trickling in coolingly; nor did he know the joy of casting a gaudy fly far ahead with a four-ounce rod, letting it drift, insect-like, over that black hole by the tree stump, and then feeling the seaweed line slip through his fingers to the whirr of the reel. And, at the end of the day, supper over, he did not squat around a big camp-fire and light his pipe, the silent darkness of the mountains gathering round, and a basketful of willow-packed trout hung in the clump of pines by the tent. Izaak's idea of fishing did not comprehend such joy. With a can of worms and a crude hook, he passed the day by quiet streams, threading the worms on his hook and thinking kindly of all things. The day's meditations over, he went back to the village, and, mayhap, joined a few kindred souls over a tankard of ale at the sign of the Red Lobster. But he missed the mountains, the water rushing past his tent, the bacon and trout, the camp-fire—the physical exaltation of it all. His kind of fishing was angling purely, while modern Waltons, as a rule, eschew the worm.
The relation of this crustacean to its host suggested to Latrobe, its discoverer, the relation of the "foretaster" in Roman times to the tyrant whom he served. A similar commensation exists in the mouth of a mullet (Mugil hospes) at Panama. The writer has received, through the courtesy of Mr. A. P. Lundin, a specimen of a flying-fish (Exonautes unicolor) taken off Sydney, Australia. To this are attached three large copepod crustaceans of the genus Penella, the largest over two inches long, and to the copepods in turn are attached a number of barnacles (Conchoderma virgatum) so joined to the copepods as to suggest strange flowers, like orchids, growing out of the fish.
Myxosporidia, or Parasitic Protozoa.—Internal parasites are very numerous and varied. Some of them are bacteria, giving rise to infectious diseases, especially in ponds and lakes. Others are myxosporidia, or parasitic protozoans, which form warty appendages, which burst, discharging the germs and leaving ulcers in their place. In the report of the U. S. Fish Commissioner for 1892, Dr. R. R. Gurley has brought together our knowledge of the protozoans of the subclass Myxosporidia, to which these epidemics are chiefly due. These creatures belong to the class of Sporozoa, and are regarded as animals, their nearest relatives being the parasitic Gregarinida, from which they differ in having the germinal portion of the spore consisting of a single protoplasmic mass instead of falciform protoplasmic rods as in the worm-like Gregarines. The Myxosporidia are parasitic on fishes, both fresh-water and marine, especially beneath the epidermis of the gills and fins and in the gall-bladder and urinary bladder. In color these protozoa are always cream-white. In size and form they vary greatly. The cyst in which they lie is filled with creamy substance made up of spores and granule matter.
"'Trout abound in the vicinity of these warm springs, presumably on account of the abundance of food there. They do not love the warm water, but usually avoid it. Several persons with whom I talked on the subject while in the park assert that diseased fish—that is to say, those which are thin and affected with flesh worms—are more commonly found near the warm water; that they take the bait readily but are logy. I frequently saw pelicans swimming near the shore in the vicinity of the warm springs on the west arm of the lake. It would appear that the badly infested or diseased fish, being less active and gamy than the healthy fish, would be more easily taken by their natural enemies, who would learn to look for them in places where they most abound. But any circumstances which cause the pelican and the trout to occupy the same neighborhood will multiply the chances of the parasites developing in both the intermediate and final host. The causes that make for the abundance of the trout parasite conspire to increase the number of adults. The two hosts react on each other and the parasite profits by the reaction. About the only enemies the trout had before tourists, ambitious to catch big strings of trout and photograph them with a kodak, began to frequent this region, were the fish-eating birds, and chief among these in numbers and voracity was the pelican. It is no wonder, therefore, that the trout should have become seriously parasitized. It may be inferred from the foregoing statements that the reason why the parasite of the trout of Yellowstone Lake migrates into the muscular tissue of its host must be found in the fact that the life of the parasite within the fish is much more prolonged than is the case where the conditions of life are less exceptional.
"The minute structure and life-history of such fungous forms have been so thoroughly made out by eminent specialists that no investigation along this line was made, save to observe those phenomena which might be easily seen with ordinary microscopic manipulations. The fungus consists of branched, hyaline filaments, without septa, except as these are found cutting off the reproductive parts of the threads. It is made up of a root-like or rhizoid part that penetrates the fish and a vegetative and reproductive part that radiates from the host. The former consists of branched tapering threads which pierce the tissues for a short distance, but are easily pulled out. The function of this part is to obtain nourishment for the growth of the external parts. Prostrate threads are found running through the natural slime covering the fish, and from these are produced the erect radiating hyphæ so plainly seen when in the water. The development of these threads appears to be very rapid when viewed under the microscope, although the growth made under favorable conditions in two days is only about a third of an inch. From actual measurements of filaments of the fungus placed in water and watched under the microscope, it was found that certain threads made a growth of about 3000 microns in an hour. Two others, watched for twenty minutes, gave in that time a growth of 90 and 47 microns respectively; and yet another filament, observed during two periods of five minutes each, made a growth of 28 microns each time. In ordinary cultures the rate of growth depends upon the condition of the medium, host, etc."
"The attack that attends the end of more lampreys than does any other is that of the fungus (Saprolegnia sp.). This looks like a gray slime and eats into the exterior parts of the animal, finally causing death. It covers the skin, the fins, the eyes, the gill-pouches, and all parts, like leprosy. It starts where the lamprey has been scratched or injured or where its mate has held it, and develops very rapidly when the water is warm. It is found late in the season on all lampreys that have spawned out, and it is almost sure to prove fatal, as we have repeatedly seen with attacked fishes or lampreys kept in tanks or aquaria. With choice aquarium fishes a remedy, or at least a palliative, is to be found in immersion in salt water for a few minutes or in bathing the affected parts with listerine. Since these creatures complete the spawning process before the fungoid attack proves serious to the individual, it can be seen that it affects no injury to the race, as the fertilized eggs are left to come to maturity. Also, as it is nature's plan that the adult lampreys die after spawning once, we are convinced that death would ensue without the attack of the fungus; and in fact this is to be regarded as a resultant of those causes that produce death rather than the immediate cause of it. Its only natural remedy is to be found in the depths of the lake (450 feet) where there is a uniform or constant temperature of about 39° Fahr., and where the light of the noon-day sun penetrates with an intensity only about equal to starlight on land on a clear but moonless night.
"In our times in Norway a sea-monster has been taken after a great storm, to which all that saw it at once gave the name of monk; for it had a man's face, rude and ungracious, the head shorn and smooth. On the shoulders, like the cloak of a monk, were two long fins instead of arms, and the end of the body was finished by a long tail. The picture I present was given me by the very illustrious lady, Margaret de Valois, Queen of Navarre, who received it from a gentleman who gave a similar one to the emperor, Charles V., then in Spain. This gentleman said that he had seen the monster as the portrait shows it in Norway, thrown by the waves and tempests on the beach at a place called Dieze, near the town called Denelopoch. I have seen a similar picture at Rome not differing in mien. Among the sea-beasts, Pliny mentions a sea-mare and a Triton as among the creatures not imaginary. Pausanias also mentions a Triton."
"This fish may be reckoned the wonder of the Ohio. It is only found as far up as the falls, and probably lives also in the Mississippi. I have seen it, but only at a distance, and have been shown some of its singular scales. Wonderful stories are related concerning this fish, but I have principally relied upon the description and picture given me by Mr. Audubon. Its length is from 4 to 10 feet. One was caught which weighed 400 pounds. It lies sometimes asleep or motionless on the surface of the water, and may be mistaken for a log or snag. It is impossible to take it in any other way than with the seine or a very strong hook; the prongs of the gig cannot pierce the scales, which are as hard as flint, and even proof against lead balls! Its flesh is not good to eat. It is a voracious fish. Its vulgar names are diamond-fish (owing to its scales being cut like diamonds), devil-fish, jackfish, garjack, etc. The snout is large, convex above, very obtuse, the eyes small and black; nostrils small, round before the eyes; mouth beneath the eyes, transversal with large angular teeth. Pectoral and abdominal fins trapezoidal. Dorsal and anal fins equal, longitudinal, with many rays. The whole body covered with large stone scales, lying in oblique rows; they are conical, pentagonal pentædral, with equal sides, from half an inch to one inch in diameter, brown at first but becoming the color of turtle-shell when dry. They strike fire with steel and are ball-proof!"
Next came Linnæus, 1758, who named Lister's figure and the species it represented, Ostracion tricornis, which should in strictness have been Ostracion tricorne, as ὀστρακίον, a little box, is a neuter diminutive. The Nagg's Head fish he named Ostracion quadricornis. The right name now is Ostracion tricornis, because the name tricornis stands first on the page in Linnæus' work, but Ostracion quadricornis has been more often used by subsequent authors because it is more truthful as a descriptive phrase. In 1798, Lacépède changed the name of Lister's fish to Ostracion listeri, a needless alteration which could only make confusion.
Ostracion cubicus is a four-angled species, and when the trunkfishes were regarded as a family (Ostraciidæ), the three-angled ones were set off as a separate genus. For this two names were offered, both by Swainson in 1839. For trigonus, a species without horns before the eyes, he gave the name Lactophrys, and for triqueter, a species without spines anywhere, the name of Rhinesomus. Most recent American authors have placed the three-cornered species which are mostly American in one genus, which must therefore be called Lactophrys. Of this name Rhinesomus is a synonym, and our species should stand as Lactophrys tricornis. The fact that Lactophrys as a word (from Latin lætus, smooth; Greek ὀφρύς, eyebrow; or else from lactoria, a milk cow, and ὀφρύς) is either meaningless or incorrectly written makes no difference with the necessity for its use.
In 1862, Bleeker undertook to divide these fishes differently. Placing all the hornless species, whether three-angled or four-angled, in Ostracion, he proposed the name Acanthostracion for the species with horns, tricornis being the type. But Acanthostracion has not been usually adopted except as the name of a section under Lactophrys. The three-angled American species are usually set apart from the four-angled species of Asia, and our cuckold is called Lactophrys tricornis. But it may be with perfect correctness called Ostracion tricorne, in the spirit called conservative. Or with the "radical" systematists we may accept the finer definition and again correctly call it Acanthostracion tricorne. But to call it quadricornis or listeri or maculatus with any generic name whatever would be to violate the law of priority.
The deep-sea work of other countries may be briefly noticed. The French vessels Travailleur and Talisman have made collections chiefly in the Mediterranean and along the coast of Africa, the results having been made known by Léon Valliant. The Hirondelle about the Azores and elsewhere has furnished material for Professor Robert Collett, of the University of Christiania. Dr. Decio Vinciguerra, of Rome, has reported on the collections of the Violante, a vessel belonging to the Prince of Monaco. Dr. A. Alcock, of Calcutta, has had charge of the most valuable deep-sea work of the Investigator in the Indian Seas. Edgar R. Waite and James Douglas Ogilby, of the Australian Museum at Sydney, have described the collections of the Thetis, on the shores of the New South Wales.
In Japan, following the scattering papers of Thunberg, Tilesius, and Houttuyn, and the monumental work of Schlegel, numerous species have been recorded by James Carson Brevoort, Günther, Gill, Eduard Nyström, Hilgendorf, and others. About 1884 Steindachner and Döderlein published the valuable "Fische Japans," based on the collections made about Tokyo by Dr. Döderlein. In 1881, Motokichi Namiye, then assistant curator in the Imperial University, published the first list of Japanese fishes by a native author. In 1900, Dr. Chiyomatsu Ishikawa, on the "Fishes of Lake Biwa," was the first Japanese author to venture to name a new species of fish (Pseudogobio zezera). This reticence was due not wholly to lack of self-confidence, but rather to the scattered condition of the literature of Japanese ichthyology. For this reason no Japanese author has ever felt that any given undetermined species was really new. Other Japanese ichthyologists of promise are Dr. Kamakichi Kishinouye, in charge of the Imperial fisheries Bureau, Dr. Shinnosuke Matsubara, director of the Imperial Fisheries Institute, Keinosuke Otaki, S. Hatta, S. Nozawa, T. Kitahara, and Michitaro Sindo, and we may look for others among the pupils of Dr. Kakichi Mitsukuri, the distinguished professor of zoology in the Imperial University.
The present writer began a "Systematic Catalogue of the Fishes of North America" in 1875 in association with his gifted friend, Herbert Edson Copeland (1849-76), whose sudden death, after a few promising beginnings, cut short the undertaking. Later, Charles Henry Gilbert (1860-), a student of Professor Copeland, took up the work and in 1883 a "Synopsis of the Fishes of North America" was completed by Jordan and Gilbert. Later, Dr. Gilbert has been engaged in studies of the fishes of Panama, Alaska, and other regions, and the second and enlarged edition of the "Synopsis" was completed in 1898, as the "Fishes of North and Middle America," in collaboration with another of the writer's students, Dr. Barton Warren Evermann. A monographic review of the Fishes of Puerto Rico was later (1900) completed by Dr. Evermann, together with numerous minor works. Other naturalists whom the writer may be proud to claim as students are Charles Leslie McKay (1854-83), drowned in Bristol Bay, Alaska, while engaged in explorations, and Charles Henry Bollman (1868-89), stricken with fever in the Okefinokee Swamps in Georgia. Still others are Dr. Carl B. Eigenmann, the indefatigable investigator of Brazilian fishes and of the blind fishes of the caves; Dr. Oliver Peebles Jenkins, the first thorough explorer of the fishes of Hawaii; Dr. Alembert Winthrop Brayton, explorer of the streams of the Great Smoky Mountains; Dr. Seth Eugene Meek, explorer of Mexico; John Otterbein Snyder, explorer of Mexico, Japan, and Hawaii; Edwin Chapin Starks, explorer of Puget Sound and Panama and investigator of fish osteology. Still other naturalists of the coming generation, students of the present writer and of his lifelong associate, Professor Gilbert, have contributed in various degrees to the present fabric of American ichthyology. Among them are Mrs. Rosa Smith Eigenmann, Dr. Joseph Swain, Wilbur Wilson Thoburn (1859-99), Frank Cramer, Alvin Seale, Albert Jefferson Woolman, Philip H. Kirsch (1860-1902), Cloudsley Rutter (died 1903), Robert Edward Snodgrass, James Francis Abbott, Arthur White Greeley, Edmund Heller, Henry Weed Fowler, Keinosuke Otaki, Michitaro Sindo, and Richard Crittenden McGregor.
The fish-remains next in age in America are from the Bloomfield sandstone in Pennsylvania of the Onondaga period in the upper Silurian. The earliest in Europe are found in the Ludlow shales, both of these localities being in or near the horizon of the Niagara rocks, in the Upper Silurian Age.
Devonian Fishes.—In the Devonian Age the Ostracophores increase in size and abundance, disappearing with the beginning of the Carboniferous. The Arthrodires also increase greatly in variety and in size, reaching their culmination in the Devonian, but not disappearing entirely until well in the Carboniferous. These two groups (often united by geologists under the older name Placoderms) together with sharks and a few Chimæras made up almost exclusively the rich fish-fauna of Devonian times. The sharks were chiefly Acanthodean and Psammodont, as far as our records show. The supposed more primitive type of Cladoselache is not known to appear before the latter part of the Devonian Age, while Pleuracanthus and Cladodus, sometimes regarded as still more primitive, are as yet found only in the Carboniferous. It is clear that the records of early shark life are still incomplete, whatever view we may adopt as to the relative rank of the different forms. Chimæroids occur in the Devonian, and with them a considerable variety of Crossopterygians and Dipnoans. The true fishes appear also in the Devonian in the guise of the Ganoid ancestors and relatives of Palæoniscum, all with diamond-shaped enameled scales. In the Devonian, too, we find the minute creature Palæospondylus, our ignorance of which is concealed under the name Cycliæ.
Mesozoic Fishes.—In the Triassic period which follows the Permian, the earliest types of Ganoids give place to forms approaching the garpike and sturgeon. The Crossopterygians rapidly decline. The Dipnoans are less varied and fewer in number; the primitive sharks, with the exception of certain Cestracionts, all disappear, only the family of Orodontidæ remaining. Here are found the first true bony fishes, doubtless derived from Ganoid stock, the allies and predecessors of the great group of herrings. Herring-like forms become more numerous in the Jurassic, and with them appear other forms which give the fish-fauna of this period something of a modern appearance. In the Jurassic the sharks become divided into several groups, Notidani, Scyllioid sharks, Lamnoid sharks, angel-fishes, skates, and finally Carcharioid sharks being now well differentiated. Chimæras are still numerous. The Acanthodei have passed away, as well as the mailed Ostrachopores and Arthrodires. The Dipnoans and Crossopterygians are few. The early Ganoids have given place to more modern types, still in great abundance and variety. This condition continues in the Cretaceous period. Here the rays and modern sharks increase in number, the Ganoids hold their own, and the other groups of soft-rayed fishes, as the smelts, the lantern-fishes, the pikes, the flying-fishes, the berycoids and the mackerels join the group of herring-like forms which represent the modern bony fishes. In the Cretaceous appear the first spiny-rayed fishes, derived probably from herring-like forms. These are allies or ancestors of the living genus Beryx.
Tertiary Fishes.—With the Eocene or first period of the Tertiary great changes have taken place. The early families of bony fishes nearly all disappear. The herring, pike, smelt, salmon, flying-fish, and berycoids remain, and a multitude of other forms seem to spring into sudden existence. Among these are the globefishes, the trigger-fishes, the catfishes, the eels, the morays, the butterfly-fishes, the porgies, the perch, the bass, the pipefishes, the trumpet-fishes, the mackerels, and the John-dories, with the sculpins, the anglers, the flounders, the blennies, and the cods. That all these groups, generalized and specialized, arose at once is impossible, although all seem to date from the Eocene times. Doubtless each of them had its origin at an earlier period, and the simultaneous appearance is related to the fact of the thorough study of the Eocene shales, which have in numerous localities (London, Monte Bolca, Licata, Mount Lebanon, Green River) been especially favorable for the preservation of these forms. Practically fossil fishes have been thoroughly studied as yet only in a very few parts of the earth. The rocks of Scotland, England, Germany, Italy, Switzerland, Syria, Ohio, and Wyoming have furnished the great bulk of all the fish remains in existence. In some regions perhaps collections will be made which will give us a more just conception of the origin of the different groups of bony fishes. We can now only say with certainty that the modern families were largely existent in the Eocene, that they sprang from ganoid stock found in the Triassic and Jurassic, that several of them were represented in the Cretaceous also, that the Berycoids were earliest of the spiny-rayed fishes, and forms allied to herring the earliest of the soft-rayed forms. Few modern families arose before the Cretaceous. Few of the modern genera go back to the Eocene, many of them arose in the Miocene, and few species have come down to us from rocks older than the end of the Pliocene. The general modern type of the fish-faunas being determined in the latter Eocene and the Miocene, the changes which bring us to recent times have largely concerned the abundance and variety of the individual species. From geological distribution we have arising the varied problems of geographical distribution and the still more complex conditions on which depend the extinction of species and of types.
The Cyclostomes.—The next group of living fishes, the Cyclostomes, including the hagfishes and lampreys,—fishes with small skull and brain but without limbs or jaws,—stands at a great distance above the lancelet in complexity of structure, and equally far from the true fishes in its primitive simplicity. In fact the lamprey is farther from the true fish in structure than a perch is from an eagle. Yet for all that it may be an offshoot from the primitive line of fish descent. There is not much in the structure of the lamprey which may be preserved in the rocks. But the cartilaginous skull, the back-bone, fins, and teeth might leave their traces in soft clay or lithographic stone. But it is certain that they have not done so in any rocks yet explored, and it may be that the few existing lampreys owe their form and structure to a process of degradation from a more complex and more fish-like ancestry. The supposed lamprey fossil of the Devonian of Scotland, Palæospondylus, has little in common with the true lampreys.
The Arthrodires.—Dimly seen in the vast darkness of Paleozoic time are the huge creatures known as Arthrodires. These are mailed and helmeted fishes, limbless so far as we know, but with sharp, notched, turtle-like jaws quite different from those of the fish or those of any animal alive to-day. These creatures appear in Silurian rocks and are especially abundant in the fossil beds of Ohio, where Newberry, Claypole, Eastman, Dean and others have patiently studied the broken fragments of their armor. Most of them have a great casque on the head with a shield at the neck and a movable joint connecting the two. Among them was almost every variation in size and form.
On the other hand, if we regard the paired fins as parts of a lateral fold of skin, we find primitive sharks to bear out our conclusions. In Cladoselache of the Upper Devonian, the pectoral and the ventral fins are long and low, and arranged just as they might be if Balfour's theory were true. Acanthoessus, with a spine in each paired fin and no other rays, might be a specialization of this type or fin, and Climatius, with rows of spines in place of pectorals and ventrals, might be held to bear out the same idea. In all these the tail is less primitive than in the Ichthyotomi. On the other hand, the vent in Cladoselache is thought by Dean to have been near the end of the tail. If this is the case, it should indicate a very primitive character. On the whole, though there is much to be said in favor of the primitive nature of the Ichthyotomi (Pleuracanthus) with the tapering tail and jointed pectoral fin of a dipnoan, and other traits of a shark, yet, on the whole, Cladoselache is probably nearer the origin of the shark-like forms.
In the second group the vertebræ have their calcareous matter arranged in rings, one or more about the notochordal center. In all these the anal fin is absent, and in the process of specialization the shark gradually gives place to the flattened body and broad fins of the ray. This group is called Tectospondyli. Those sharks of this group with one ring of calcareous matter in each vertebra constitute the most primitive extreme of a group representing continuous evolution.
The Chimæras.—The most ancient offshoot from the shark stem, perhaps dating from Silurian times and possibly separated at a period earlier than the date of any known shark, is the group of Holocephali or Chimæras, shark-like in essentials, but differing widely in details. Of these there are but few living forms and the fossil types are known only from dental plates and fin-spines. The living forms are found in the deeper seas the world over, one of the simplest in structure being the newly discovered Rhinochimæra of Japan. The fusion of the teeth into overlapping plates, the covering of the gills by a dermal flap, the complete union of the palato-quadrate apparatus or upper jaw with the skull and the development of a peculiar clasping spine on the forehead of the male are characteristic of the Chimæras. The group is one of the most ancient, but it ends with itself, none of the modern fishes being derived from Chimæras.
The Crossopterygians.—The fish-like mouth appears with the group of Crossopterygians, fishes which still retain the old-fashioned type of pectoral and ventral fin, the archipterygium. In the archaic tail, enameled scales, and cartilaginous skeleton the Crossopterygian shows its affinity with its Dipnoan ancestry. Thus these fishes unite in themselves traits of the shark, lung-fish, and Ganoid. The few living Crossopterygians, Polypterus and Erpetoichthys, are not very different from those which prevailed in Devonian times. The larvæ possess external gills with firm base and fringe-like rays, suggesting a resemblance to the pectoral fin itself, which develops from the shoulder-girdle just below it and would seem to give some force to Kerr's contention that the archipterygium is only a modified external gill. In Polypterus the archipterygium has become short and fan-shaped, its axis made of two diverging bones with flat cartilage between. From this type it is thought that the arm of the higher forms has been developed. The bony basis may be the humerus, from which diverge radius and ulna, the carpal bones being formed of the intervening cartilage.
The Actinopteri.—From the Crossopterygians springs the main branch of the true fishes, known collectively as Actinopteri, or ray-fins, those with ordinary rays on the paired fins instead of the jointed archipterygium. The transitional series of primitive Actinopteri are usually known as Ganoids. The Ganoid differs from the Crossopterygian in having the basal elements of the paired fins small and concealed within the flesh. But other associated characters of the Crossopterygii and Dipnoans are preserved in most of the species. Among these are the mailed head and body, the heterocercal tail, the cellular air-bladder, the presence of valves in the arterial bulb, the presence of a spiral valve in the intestine and of a chiasma in the optic nerves. All these characters are found in the earlier types so far as is known, and all are more or less completely lost or altered in the teleosts or bony fishes. Among these early types is every variety of form, some of them being almost as long as deep, others arrow-shaped, and every intermediate form being represented. An offshoot from this line is the bowfin (Amia calva), among the Ganoids the closest living ally of the bony fishes, showing distinct affinities with the great group to which the herring and salmon belong. Near relatives of the bowfin flourished in the Mesozoic, among them some with a forked tail, and some with a very long one. From Ganoids of this type the vast majority of recent fishes may be descended.
From the abdominal forms allied to the herring, the huge array of modern fishes, typified by the perch, the bass, the mackerel, the wrasse, the globefish, the sculpin, the sea-horse, and the cod descended in many diverging lines. The earliest of the spine-rayed fishes with thoracic fins belong to the type of Berycidæ, a group characterized by rough scales, the retention of a primitive bone between the eyes, and the retention of the primitive larger number of ventral rays. These appear in the Cretaceous or chalk deposits, and show various attributes of transition from the abdominal to the thoracic type of ventrals.
Another line of descent apparently distinct from that of the herring and salmon extends through the characins to the loach, carps, catfishes, and electric eel. The fishes of this series have the anterior vertebræ coossified and modified in connection with the hearing organ, a structure not appearing elsewhere among fishes. This group includes the majority of fresh-water fishes. Still another great group, the eels, have lost the ventral fins and the bones of the head have suffered much degradation.
The most highly developed fishes, all things considered, are doubtless the allies of the perch, bass, and sculpin. These fishes have lost the air-duct and on the whole they show the greatest development of the greatest number of structures. In these groups their traits one after another are carried to an extreme and these stages of extreme specialization give way one after another to phases of degeneration. The specialization of one organ usually involves degeneration of some other. Extreme specialization of any organ tends to render it useless under other conditions and may be one step toward its final degradation.
We have thus seen, in hasty review, that the fish-like vertebrates spring from an unknown and possibly worm-like stock, that from this stock, before it became vertebrate, degenerate branches have fallen off, represented to-day by the Tunicates and Enteropneustans. We have seen that the primitive vertebrate was headless and limbless and without hard parts. The lancelet remains as a possible direct offshoot from it; the cyclostome with brain and skull is a possible derivative from archaic lancelets. The earliest fishes leaving traces in the rocks were mailed ostracophores. From an unknown but possibly lamprey-like stock sprang the sharks and chimæras. The sharks developed into rays in one right line and into the highest sharks along another, while by a side branch through lost stages the primitive sharks passed into Crossopterygians, into Dipnoans, or lung-fishes, and perhaps into Ostracophores. All these types and others abound in the Devonian Age and the early records were lost in the Silurian. From the Crossopterygians or their ancestors or descendants by the specialization of the lung and limbs, the land animals, at first amphibians, after these reptiles, birds, and mammals, arose.
The Enteropneusta.—Most simple, most worm-like, and perhaps most primitive of all the Chordates is the group of worm-shaped forms, forming the class of Enteropneusta. The class of Enteropneusta, also called Adelochorda or Hemichordata, as here recognized, consists of a group of small marine animals allied to the genus Balanoglossus, or acorn-tongues (βάλανος, acorn; γλώσσα, tongue). These are worm-like creatures with fragile bodies buried in the sand or mud, or living under rocks of the seashore and in shallow waters, where they lie coiled in a spiral, with little or no motion. From the surface of the body a mucous substance is secreted, holding together particles by which are formed tubes of sand. The animal has a peculiar odor like that of iodoform. At the front is a long muscular proboscis, very sensitive, capable of great extension and contraction, largely used in burrowing in the ground, and of a brilliant orange color in life. Behind this is a collar which overlaps the small neck and conceals the small mouth at the base of the proboscis. The gill-slits behind the collar are also more or less concealed by it.
A peculiar little creature, known as Tornaria, was once considered to be the larva of a starfish. It is minute and transparent, floating on the surface of the sea. It has no visible resemblance to the adult Balanoglossus, but it has been reared in aquaria and shown to pass into the latter or into the related genus Glossobalanus. No such metamorphosis was found by Bateson in the more primitive genus Dolichoglossus, studied by him. This adult animal may be, indeed, a worm as it appears, but the presence of gill-slits, the existence of a rudimentary notochord, and the character of the central nervous system are distinctly fish-like and therefore vertebrate characters. With the Chordates, and not with the worms, this class, Enteropneusta (ἔντερον, intestine; πνεῖν, to breathe), must be placed if its characters have been rightly interpreted. It is possibly a descendant of the primitive creatures which marked the transition from the archaic worms, or possibly archaic Echinoderms, to the archaic Chordate type.
Classification of Enteropneusta.—Until recently the Enteropneusta have been usually placed in a single family or even in a single genus. The recent researches of Professor J. W. Spengel of Giessen and of Professor William Emerson Ritter of the University of California, have shown clearly that the group is much larger than had been generally supposed, with numerous species in all the warm seas. In Spengel's recent paper, "Die Benennung der Enteropneusten-Gattungen," three families are recognized with nine genera and numerous species. At least seven species are now known from the Pacific Coast of North America.
Development of Tunicates.—The sexes are combined in the same individual, though usually the products ripen at different times. As a rule, the earlier stages of the embryo are passed inside the cloacal chamber, though in some the development occurs outside the body. As a type of the development we will consider that of one of the solitary forms, leaving the many curious modifications to be noticed in connection with the species in which they occur. This will be best, since these forms show the relationship to the other vertebrates in the clearest manner.
The egg undergoes a total segmentation and a regular gastrulation. Soon a tail appears, and under the microscope the young embryo, which now begins its free life, appears much like the tadpole of the frog. It has a large oval body and a long tail which lashes about, forcing the animal forward with a wriggling motion. Nor is the resemblance superficial; it pervades every part of the structure, as may be seen from the adjacent diagram. The mouth is nearly terminal and communicates with a gill-chamber provided with gill-clefts. At the posterior end of the gill-chamber begins the alimentary tract, which pursues a convoluted course to the vent. In the tail, but not extending to any distance into the body, is an axial cylinder, the notochord, which here, as in all other vertebrates, arises from the hypoblast; and above it is the spinal cord (epiblastic in origin), which extends forward to the brain, above the gill-chamber. Besides, the animal is provided with organs of sight and hearing, which, however, are of peculiar construction and can hardly be homologized with the corresponding organs in vertebrates. So far the correspondence between the two types is very close, and if we knew nothing about the later stages, one would without doubt predict that the adult tunicate would reach a high point in the scale of vertebrates. These high expectations are never fulfilled; the animal, on the contrary, pursues a retrograde course, resulting in an adult whose relationship to the true vertebrates never would have been suspected had its embryology remained unknown.
Larvacea.—In the most primitive order the animals are minute and free-swimming, never passing beyond the tadpole stage. The notochord and the nervous chord persist through life, the latter with ganglionic segmentations at regular intervals. The species mostly float in the open sea, and some of them form from their own secretions a transparent gelatinous envelope called a "house." This has two apertures and a long chamber "in which the tail has room to vibrate."
In the Botryllidæ and Polystyelidæ the individuals are not segmented and in the former family are arranged in star-shaped groups about a common cloaca, into which the atrial siphons of the different individuals open. The group springs by budding from the tadpole, or larva, which has attached itself to some object. These forms are often brightly colored. Botryllus gouldi is a species very common along our North Atlantic coast, forming gray star-shaped masses sometimes an inch across on eel-grass (Zostera) and on flat-leaved seaweeds. Goodsiria dura, a representative of the Polystyelidæ, is one of the most common Ascidians on the California coast southward, where the brick-red masses incrusting on seaweeds of various kinds, and on other Ascidians, are frequently thrown ashore in great quantities during heavy storms.
The Salpas abound in the warm seas, the chains often covering the water for miles. They are perfectly transparent, and the chains are often more than a foot in length. In Doliolum the body is barrel-shaped and the gills are less modified than in Salpa. The alternation of generations in this genus is still more complicated than in Salpa, for here we have not only a sexual and a non-sexual generation, the individuals of which differ from each other, but there is further a differentiation among the asexually produced individuals themselves; so that we have in all three instead of two sorts of animals in the complete life cycle. Besides the proliferating stolon situated on the ventral side, the bud-producing individual possesses a dorsal process larger than the stolon proper. The buds become completely severed from the true stolon at an early stage and actually crawl along the side of the parent up to the dorsal process, upon which they arrange themselves in three rows, two lateral and one median. The buds of the lateral rows become nutritive and respiratory zooids, while those of the median row, ultimately at least, give rise in turn to the egg-producing individuals.
Species of Lancelets.—The Mediterranean species ranges northward to the south of England. Others are found as far north as Chesapeake Bay, San Diego, and Misaki in Japan, where is found a species called Branchiostoma belcheri. The sands at the mouth of San Diego Bay are noted as producing the largest of the species of lancelets, Branchiostoma californiense. From the Bahamas comes the smallest, the type of a distinct genus, Asymmetron lucayanum, distinguished among other things by a projecting tail. Other supposed genera are Amphioxides (pelagicus), dredged in the deep sea off Hawaii and supposed to be pelagic, the mouth without cirri; Epigonichthys (cultellus), from the East Indies, and Heteropleuron (bassanum), from Bass Straits, Australia. These little animals are of great interest to anatomists as giving the clue to the primitive structure of vertebrates. While possibly these have diverged widely from their actual common ancestry with the fishes, they must approach near to these in many ways. Their simplicity is largely primitive, not, as in the Tunicates, the result of subsequent degradation.
Origin of Lancelets.—It is doubtless true, as Dr. Willey suggests, that the Vertebrates became separated from their worm-like ancestry through "the concentration of the central nervous system along the dorsal side of the body and its conversion into a hollow tube." Besides this trait two others are common to all of them, the presence of the gill-slits and that of the notochord. The gill-slits may have served primarily to relieve the stomach of water, as in the lowest forms they enter directly into the body-cavity. The primitive function of the notochord is still far from clear, but its ultimate use of its structures in affording protection and in furnishing a fulcrum for the muscles and limbs is of the greatest importance in the processes of life.
Conodontes.—Certain structures found as fossils have been from time to time regarded as Cyclostomes, but in all such cases there is doubt as to the real nature of the fossil relic in question or as to the proper interpretation of its relationship.
The Hyperotreta, or Hagfishes.—The Hyperotreta (ὑπερῴα, palate; τρετός, perforate), or hagfishes, have the nostril highly developed, a tube-like cylinder with cartilaginous rings penetrating the palate. In these the eyes are little developed and the species are parasitic on other fishes. In Polistotrema stouti, the hagfish of the coast of California, is parasitic on large fishes, rockfishes, or flounders. It usually fastens itself at the throat or isthmus of its host and sometimes at the eyes. Thence it works very rapidly to the inside of the body. It there devours all the muscular part of the fish without breaking the skin or the peritoneum, leaving the fish a living hulk of head, skin, and bones. It is especially destructive to fishes taken in gill-nets. The voracity of the Chilean species Polistotrema dombeyi is equally remarkable. Dr. Federico T. Delfin finds that in seven hours a hagfish of this species will devour eighteen times its own weight of fish-flesh. The intestinal canal is a simple tube, through which most of the food passes undigested. The eggs are large, each in a yellowish horny case, at one end of which are barbed threads by which they cling together and to kelp or other objects. In the California hagfish, Polistotrema stouti, great numbers of these eggs have been found in the stomachs of the males.
The lampreys of the northern regions constitute the family of Petromyzonidæ. The larger species (Petromyzon, Entosphenus) live in the sea, ascending rivers to spawn, and often becoming land-locked and reduced in size by living in rivers only. Such land-locked marine lampreys (Petromyzon marinus unicolor) breed in Cayuga Lake and other lakes in New York. The marine forms reach a length of three feet. Smaller lampreys of other genera six inches to eighteen inches in length remain all their lives in the rivers, ascending the little brooks in the spring, clinging to stones and clods of earth till their eggs are deposited. These are found throughout northern Europe, northern Asia, and the colder parts of North America, belonging to the genera Lampetra and Ichthyomyzon. Other and more aberrant genera from Chile and Australia are Geotria and Mordacia, the latter forming a distinct family, Mordaciidæ. In Geotria, a large and peculiar gular pouch is developed at the throat. In Macrophthalmia chilensis from Chile the eyes are large and conspicuous.
Metamorphosis of Lampreys.—All lampreys, so far as known, pass through a distinct metamorphosis. The young, known as the Ammocœtes form, are slender, eyeless, and with the mouth narrow and toothless. From Professor Surface's paper on "The Removal of Lampreys from the Interior Waters of New York" we have the following extracts (slightly condensed):
Migration or "Running" of Lampreys.—"After thus feeding to an unusual extent, their reproductive elements (gonads) become mature and their alimentary canals commence to atrophy. This duct finally becomes so occluded that from formerly being large enough to admit a lead-pencil of average size when forced through it, later not even liquids can pass through, and it becomes nearly a thread closely surrounded by the crowding reproductive organs. When these changes commence to ensue, the lampreys turn their heads against the current and set out on their long journeys to the sites that are favorable for spawning, which here may be from two to eight miles from the lake. In this migration they are true to their instincts and habits of laziness in being carried about, as they make use of any available object, such as a fish, boat, etc., that is going in their direction, fastening to it with their suctorial mouths and being borne along at their ease. During this season it is not infrequent that as the Cornell crews come in from practice and lift their shells from the water, they find lampreys clinging to the bottoms of the boats, sometimes as many as fifty at one time. They are likely to crowd up all streams flowing into the lake, inspecting the bed of the stream as they go. They do not stop until they reach favorable spawning sites, and if they find unsurmountable obstacles in their way, such as vertical falls or dams, they turn around and go down-stream until they find another, up which they go. This is proved every spring by the number of adult lampreys which are seen temporarily in Pall Creek and Cascadilla Creek. In each of these streams, about a mile from its mouth, there is a vertical fall over thirty feet in height which the lampreys cannot surmount, and in fact they have never been seen attempting to do so. After clinging with their mouths to the stones at the foot of the falls for a few days, they work their way down-stream, carefully inspecting all the bottom for suitable spawning sites. They do not spawn in these streams because there are too many rocks and no sand, but finally enter the only stream (the Cayuga Lake inlet) in which they find suitable and accessible spawning sites.
What becomes of Lampreys after Spawning?—"There has been much conjecture as to the final end of the lampreys, some writers contending that they die after spawning, others that they return to deep water and recuperate, and yet others compromise these two widely divergent views by saying that some die and others do not. The fact is that the spawning process completely wears out the lampreys, and leaves them in a physical condition from which they could never recover. They become stone-blind; the alimentary canal suffers complete atrophy; their flesh becomes very green from the katabolic products, which find the natural outlet occluded; they lose their rich yellow color and plump, symmetrical appearance; their skin becomes torn, scratched, and worn off in many places, so that they are covered with sores, and they become covered with a parasitic or sarcophytic fungus, which forms a dense mat over almost their entire bodies, and they are so completely debilitated and worn out that recovery is entirely out of the question. What is more, the most careful microscopical examination of ovaries and testes has failed to reveal any evidence of new gonads or reproductive bodies. This is proof that reproduction could not again ensue without a practical rebuilding of the animals, even though they should regain their vitality. A. Mueller, in 1865, showed that all the ova in the lamprey were of the same size, and that after spawning no small reproductive bodies remained to be developed later. This is strong evidence of death after once spawning.
Hasse's Classification of Elasmobranchs.—In 1882, Professor Carl Hasse proposed to subdivide the sharks on the basis of the structure of the individual vertebræ. In the lowest group, a hypothetical order of Polyospondyli, possibly represented by the fossil spines called Onchus, an undivided notochord, perhaps swollen at regular intervals, is assumed to have represented the vertebral column. In the Diplospondyli (Hexanchidæ) the imperfectly segmented vertebræ are joined in pairs, each pair having two neural arches. In the Asterospondyli or ordinary sharks each vertebra has its calcareous lamella radiating star-like from the central axis. In the Cyclospondyli (Squalidæ, etc.) the calcareous part forms a single ring about the axis, and in the Tectospondyli (Squatina, rays, etc.) it forms several rings. These groups again are natural and correspond fairly with those based on other characters. At the same time there is no far-reaching difference between Cyclospondyli and Tectospondyli, and the last-named section includes both sharks and rays.
For our purpose, without taking sides in this controversy, we may follow Dean in allowing Cladoselache to stand as the most primitive of known sharks, thus arranging the Elasmobranchs and rays, recent and fossil, in six orders of unequal value—Pleuropterygii, Acanthodei, Ichthyotomi, Notidani, Asterospondyli, and Tectospondyli. Of these orders the first and second are closely related, as are also the fourth and fifth, the sixth being not far remote. The true sharks form the culmination of one series, the rays of another, while from the Ichthyotomi the Crossopterygians and their descendants may be descended. But this again is very hypothetical, or perhaps impossible; while, on the other hand, the relation of the Chimæras to the sharks is still far from clearly understood.
Order Acanthodei.—Near the Pleuropterygii, although much more highly developed, we may note the strange group of Acanthodei (ἀκανθώδης, spinous). These armed fishes were once placed among the Crossopterygians, but there seems no doubt that Woodward is right in regarding them as a highly specialized aberrant offshoot of the primitive sharks. In this group the paired fins consist each of a single stout spine, nearly or quite destitute of other rays. A similar spine is placed in front of the dorsal fin and one in front of the anal. According to Dean these spines are each produced by the growing together of all the fin-rays normally belonging to the fin, a view of their morphology not universally accepted.
The species of Acanthoessus are all small fishes rarely more than a foot long, with very small teeth or none, and with the skin well armed with a coat-of-mail. Acanthoessus bronni is the one longest known. In the earliest species known, from the Devonian, the ventral fins are almost as large as the pectorals and nearly midway between pectorals and anal. In the later species the pectoral fins become gradually larger and the ventrals move forward. In the Permian species the pectorals are enormous.
Dean on Acanthodei.—In his latest treatise on these fishes, "The Devonian Lamprey," Dr. Dean unites the Pleuropterygii and Acanthodei in a single order under the former name, regarding Acanthoessus as an ally and perhaps descendant of the primitive Cladoselache. Dr. Dean observes:
From these primitive extinct types of shark we may proceed to those forms which have representatives among living fishes. From Cladoselache a fairly direct series extends through the Notidani and Cestraciontes, culminating in the Lamnoid and Galeoid sharks.
Order Ichthyotomi.—In the order Ichthyotomi (ἰχθύς, fish; τομός, cutting; named by Cope from the supposed segmentation of the cranium; called by Parker and Haswell Pleuracanthea) the very large pectoral fins are developed each as an archipterygium. Each fin consists of a long segmented axis fringed on one or both sides with fin-rays. The notochord is very simple, scarcely or never constricted, the calcifications of its sheath "arrested at the most primitive or rhachitomous stage, except in the tail." This is the best defined of the orders of sharks, and should perhaps rank rather as a subclass, as the Holocephali. Two families of Ichthyotomi are recognized by Woodward, the Pleuracanthidæ and the Cladodontidæ. In the Pleuracanthidæ the dorsal fin is long and low, continuous from head to tail, and the pectoral rays are in two rows. There is a long barbed spine with two rows of serrations at the nape. The body is slender, not depressed, and probably covered with smooth skin. The teeth have two or more blunt cusps, sometimes with a smaller one between and a blunt button behind. The interneural cartilages are more numerous than the neural spines. The genera are imperfectly known, the skeleton of Pleuracanthus decheni only being well preserved. This is the type of the genus called Xenacanthus which, according to Woodward, is identical with Pleuracanthus, a genus otherwise known from spines only. The denticles on the spine are straight or hooked backward, in Pleuracanthus (lævissimus), the spine being flattened. In Orthacanthus (cylindricus), the spine is cylindrical in section. The species called Dittodus and Didymodus are known from the teeth only. These resemble the teeth of Chlamydoselachus. It is not known that Dittodus possesses the nuchal spine, although detached spines like those of Pleuracanthus lie about in remains called Didymodus in the Permian rocks of Texas. In Dicranodus texensis the palato-quadrate articulates with the postorbital process of the cranium, as in the Hexanchidæ, and the hyomandibular is slender.
A genus, Chondrenchelys, from the sub-Carboniferous of Scotland, is supposed to belong to the Pleuracanthidæ, from the resemblance of the skeleton. It has no nuchal spine, and no trace of paired fins is preserved.
Fossil Hexanchidæ exist in large numbers, all of them referred by Woodward to the genus Notidanus (which is a later name than Hexanchus and Heptranchias and intended to include both these genera), differing chiefly in the number of gill-openings, a character not ascertainable in the fossils. None of these, however, appear before Cretaceous time, a fact which may indicate that the simplicity of structure in Hexanchus and Heptranchias is a result of degeneration and not altogether a mark of primitive simplicity. The group is apparently much younger than the Cestraciontes and little older than the Lamnoids, or the Squaloid groups. Heptranchias microdon is common in English Cretaceous rocks, and Heptranchias primigenius and other species are found in the Eocene.
Order Asterospondyli.—The order of Asterospondyli comprises the typical sharks, those in which the individual vertebræ are well developed, the calcareous lamellæ arranged so as to radiate, star-fashion, from the central axis. All these sharks possess two dorsal fins and one anal fin, the pectoral fin is normally developed, with the three basal cartilages; there are five gill-openings, and the tail is heterocercal.
These forms find their nearest allies in the Notidani, which they resemble to some extent in dentition and in having the palato-quadrate articulated to the skull although fastened farther forward than in the Notidani. Each of the two dorsal fins has a strong spine.
Family Heterodontidæ.—Among recent species this group contains only the family of Heterodontidæ, the bullhead sharks, or Port Jackson sharks. In this family the head is high, with usually projecting eyebrows, the lateral teeth are pad-like, ridged or rounded, arranged in many rows, different from the pointed anterior teeth, the fins are large, the coloration is strongly marked, and the large egg-cases are spirally twisted. All have five gill-openings. The living species of Heterodontidæ are found only in the Pacific, the Port Jackson shark of Australia, Heterodontus philippi, being longest known. Other species are Heterodontus francisci, common in California, Heterodontus japonicus, in Japan, and Heterodontus zebra, in China. These small and harmless sharks at once attract attention by their peculiar forms. In the American species the jaws are less contracted than in the Asiatic species, called Heterodontus. For this reason Dr. Gill has separated the former under the name of Gyropleurodus. The differences are, however, of slight value. The genus Heterodontus first appears in the Jurassic, where a number of species are known, one of the earliest being Heterodontus falcifer.
These remarkable Ichthyodorulites are characteristic structures of sharks of unknown nature, but probably related to the Heterodontidæ. Of these the principal genera are Edestus, Helicoprion, and Campyloprion. Karpinsky regards these ornate serrated spiral structures as whorls of unshed teeth cemented together and extending outside the mouth, "sharp, piercing teeth which were never shed but became fused in whorls as the animals grew."
To a hypothetical group of primitive sharks Dr. Hasse has given the name of Polyospondyli. In these supposed ancestral sharks the vertebræ were without any ossification, a simple notochord, possibly swollen at intervals. The dorsal fin was single and long, a fold of skin with perhaps a single spine as an anterior support. The teeth must have been modified dermal papillæ, each probably with many cusps. Probably seven gill-openings were developed, and the tail was diphycercal, ending in a straight point. The finely striated fin-spines not curved upward at tip, called Onchus from the Upper Silurian of the Ludlow shales of England and elsewhere, are placed by Hasse near his Polyspondylous sharks. Such spines have been retained by the group of Chimæras, supposed to be derived from the ancestors of Onchus, as well as by the Heterodontidæ and Squalidæ.
The goblin-shark, or Tenguzame, Mitsukurina owstoni, is a very large shark rarely taken in the Kuro Shiwo, or warm "Black Current" of Japan. It is characterized by the development of the snout into a long flat blade, extending far beyond the mouth, much as in Polyodon and in certain Chimæras. Several specimens are now known, all taken by Capt. Alan Owston of Yokohama in Sagami Bay, Japan. The original specimen, a young shark just born, was presented by him to Professor Kakichi Mitsukuri of the University of Tokyo. From this our figure was taken. The largest specimen now known is in the United States National Museum and is fourteen feet in length. In the Upper Cretaceous is a very similar genus, Scapanorhynchus (lewisi, etc.), which Professor Woodward thinks may be even generically identical with Mitsukurina, though there is considerable difference in the form of the still longer rostral plate, and the species of Scapanorhynchus differ among themselves in this regard.
Family Pseudotriakidæ.—The Pseudotriakidæ consist of two species. One of these is Pseudotriakis microdon, a large shark with a long low tail, long and low dorsal fin, and small teeth. It has been only twice taken, off Portugal and off Long Island. The other, the mute shark, Pseudotriakis acrales, a large shark with the body as soft as a rag, is in the museum of Stanford University, having been taken by Mr. Owston off Misaki.
In the closely related genus, Isurus, the mackerel-sharks, this cusp is wanting, while in Isuropsis the dorsal fin is set farther back. In each of these genera the species reach a length of 20 to 25 feet. Each is strong, swift, and voracious. Isurus oxyrhynchus occurs in the Mediterranean, Isuropsis dekayi, in the Gulf of Mexico, and Isuropsis glauca, from Hawaii and Japan westward to the Red Sea.
Carcharodon mega odon.
539 Cetorhinus maximus, Basking-shark.
540 Galeus zyopterus, Soup-fin Shark.
541 Carcharias lamia, Cub-shark.
542Teeth of
Corax pristodontus.
543 Sphyrna zygæna, Hammer-head Shark.
544 Squalus acanthias, Dogfish.
545Etmopterus lucifer.
546Brain of Monkfish,
Squatina squatina.
547 Pristiophorus japonicus, Saw-shark.
548 Pristis pectinatus, Sawfish.
550 Rhinobatus lentiginosus, Guitar-fish.
551 Raja erinacea, Common Skate.
552 Narcine brasiliensis, Numbfish.
553Teeth of
Janassa linguæformis.
554Polyrhizodus radicans.
555 Dasyatis sabina, Sting-ray.
556 Aëtobatis narinari, Eagle-ray.
558 Manta birostris, Devil-ray or Sea-devil.
559Skeleton of
Chimæra monstrosa.
564 Chimæra colliei, Elephant-fish.
565 Odontotodus schrencki, Ventral Side.
570 Odontotodus schrencki, Dorsal Side.
570Head of
Odontotodus schrencki, from the Side.
571 Limulus polyphemus, Horseshoe Crab.
572Lanarkia spinosa.
574 [Pg xxviii] Drepanaspis gmundenensis. 575Pteraspis rostrata.
575 Cephalaspis lyelli, Restored.
576Cephalaspis dawsoni.
577Pterichthyodes testudinarius.
578 Pterichthyodes testudinarius, Side View.
579Birkenia elegans.
579Lasianius problematicus.
580 Coccosteus cuspidatus, Restored.
582Jaws of
Dinichthys hertzeri.
583 Dinichthys intermedius, an Arthrodire.
584Palæospondylus gunni.
591Shoulder-girdle of
Polypterus bichir.
600Arm of a Frog.
601 Polypterus congicus, a Crossopterygian Fish.
602Basal Bone of Dorsal Fin,
Holoptychius leptopterus.
603Gyroptychius microlepidotus.
604 Cœlacanthus elegans, showing Air-bladder.
604Undina gulo.
605Lower Jaw of
Polypterus bichir, from Below.
606Polypterus congicus.
607Polypterus delhezi.
607Erpetoichthys calabaricus.
608Shoulder-girdle of
Neoceratodus forsteri.
609Phaneropleuron andersoni.
613Teeth of
Ceratodus runcinatus.
614Neoceratodus forsteri.
614Archipterygium of
Neoceratodus forsteri.
614Upper jaw of
Neoceratodus forsteri.
615Lower Jaw of
Neoceratodus forsteri.
616Adult Male of
Lepidosiren paradoxa.
619 Lepidosiren paradoxa.Embryo Three Days before Hatching; Larva Thirteen Days after Hatching.
620Larva of
Lepidosiren paradoxaForty Days after Hatching.
621Larva of
Lepidosiren paradoxaThirty Days after Hatching.
621Larva of
Lepidosiren paradoxaThree Months after Hatching.
621Protopterus dolloi.
622Family Cetorhinidæ, or Basking Sharks.—The largest of all living sharks is the great basking shark (Cetorhinus maximus), constituting the family of Cetorhinidæ. This is the largest of all fishes, reaching a length of thirty-six feet and an enormous weight. It is a dull and sluggish animal of the northern seas, almost as inert as a sawlog, often floating slowly southward in pairs in the spring and caught occasionally by whalers for its liver. When caught, its huge flabby head spreads out wide on the ground, its weight in connection with the great size of the mouth-cavity rendering it shapeless. Although so clumsy and without spirit, it is said that a blow with its tail will crush an ordinary whaleboat. The basking shark is known on all northern coasts, but has most frequently been taken in the North Sea, and about Monterey Bay in California. From this locality specimens have been sent to the chief museums of Europe. In its external characters the basking shark has much in common with the man-eater. Its body is, however, relatively clumsy forward; its fins are lower, and its gill-openings are much broader, almost meeting under the throat. The great difference lies in the teeth, which in Cetorhinus are very small and weak, about 200 in each row. The basking shark, also called elephant-shark and bone-shark, does not pursue its prey, but feeds on small creatures to be taken without effort. Fossil teeth of Cetorhinus have been found from the Cretaceous, as also fossil gill-rakers, structures which in this shark are so long as to suggest whalebone.
The Carcharioid Sharks, or Requins.—The largest family of recent sharks is that of Carchariidæ (often called Galeorhinidæ, or Galeidæ), a modern offshoot from the Lamnoid type, and especially characterized by the presence of a third eyelid, the nictitating membrane, which can be drawn across the eye from below. The heterocercal tail has no keel; the end is bent upward; both dorsal fins are present, and the first is well in front of the ventral fins; the last gill-opening over the base of the pectoral, the head normally formed; these sharks are ovoviviparous, the young being hatched in a sort of uterus, with or without placental attachment.
Galeocerdo differs mainly in having a pit at the root of the tail. Its species, large, voracious, and tiger-spotted, are found in warm seas and known as tiger-sharks (Galeocerdo maculatus in the Atlantic, Galeocerdo tigrinus in the Pacific).
Probably allied to the Carchariidæ is the genus Corax, containing large extinct sharks of the Cretaceous with broadtriangular serrate teeth, very massive in substance, and without denticles. As only the teeth are known, the actual relations of the several species of Corax are not certainly known, and they may belong to the Lamnidæ.
The Order of Tectospondyli.—The sharks and rays having no anal fin and with the calcareous lamellæ arranged in one or more rings around a central axis constitute a natural group to which, following Woodward, we may apply the name of Tectospondyli. The Cyclospondyli (Squalidæ, etc.) with one ring only of calcareous lamellæ may be included in this order, as also the rays, which have tectospondylous vertebræ and differ from the sharks as a group only in having the gill-openings relegated to the lower side by the expansion of the pectoral fins. The group of rays and Hasse's order of Cyclospondyli we may consider each as a suborder of Tectospondyli. The origin of this group is probably to be found in or near the Cestraciontes, as the strong dorsal spines of the Squalidæ resemble those of the Heterodontidæ.
The genus Squalus includes the dogfishes, small, greedy sharks abundant in almost all cool seas and in some tropical waters. They are known by the stout spines in the dorsal fins and by their sharp, squarish cutting teeth. They are largely sought by fishermen for the oil in their livers, which is used to adulterate better oils. Sometimes 20,000 have been taken in one haul of the net. They are very destructive to herrings and other food-fishes. Usually the fishermen cut out the liver, throwing the shark overboard to die or to be cast on the beach. In northern Europe and New England Squalus acanthias is abundant. Squalus sucklii replaces it in the waters about Puget Sound, and Squalus mitsukurii in Japan and Hawaii. Still others are found in Chile and Australia. The species of Squalus live near shore and have the gray color usual among sharks. Allied forms perhaps hardly different from Squalus are found in the Cretaceous rocks and have been described as Centrophoroides. Other genera related to Squalus live in greater depths, from 100 to 600 fathoms, and these are violet-black. Some of the deep-water forms are the smallest of all sharks, scarcely exceeding a foot in length. Etmopterus spinax lives in the Mediterranean, and teeth of a similar species occur in the Italian Pliocene rocks. Etmopterus lucifer,[150] a deep-water species of Japan, has a brilliant luminous glandular area along the sides of the belly. Other small species of deeper waters belong to the genera Centrophorus, Centroscymnus, and Deania. In some of these species the scales are highly specialized, pedunculate, or having the form of serrated leaves. Some species are Arctic, the others are most abundant about Misaki in Japan and the Madeira Islands, two regions especially rich in semi-bathybial types. Allied to the Squalidæ is the small family of Oxynotidæ with short bodies and strong dorsal spine. Oxynotus centrina is found in the Mediterranean, and its teeth occur in the Miocene.
The principal family is that of the angel-fishes, or monkfishes (Squatinidæ). In this group the body is depressed and flat like that of a ray. The greatly enlarged pectorals form a sort of shoulder in front alongside of the gill-openings, which has suggested the bend of the angel's wing. The dorsals are small and far back, the tail is slender with small fins, all these being characters shared by the rays. But one genus is now extant, widely diffused in warm seas. The species if really distinct are all very close to the European Squatina squatina. This is a moderate-sized shark of sluggish habit feeding on crabs and shells, which it crushes with its small, pointed, nail-shaped teeth. Numerous fossil species of Squatina are found from the Triassic and Cretaceous, Squatina alifera being the best known.
Pristididæ, or Sawfishes.—The sawfishes, Pristididæ, are long, shark-like rays of large size, having, like the sawsharks, the snout prolonged into a very long and strong flat blade, with a series of strong enameled teeth implanted in sockets along either side of it. These teeth are much larger and much less sharp than in the sawsharks, but they are certainly homologous with these, and the two groups must have a common descent, distinct from that of the other rays. Doubtless when taxonomy is a more refined art they will constitute a small suborder together. This character of enameled teeth on the snout would seem of more importance than the position of the gill-openings or even the flattening and expansion of the body. The true teeth in the sawfishes are blunt and close-set, pavement-like as befitting a ray. (See Fig. 152.)
Rhinobatidæ, or Guitar-fishes.—The Rhinobatidæ (guitar-fishes) are long-bodied, shovel-nosed rays, with strong tails; they are ovoviviparous, hatching the eggs within the body. The body, like that of the shark or sawfish, is covered with nearly uniform shagreen. The numerous species abound in all warm seas; they are olive-gray in color and feed on small animals of the seabottoms. The length of the snout differs considerably in different species, but in all the body is relatively long and strong. Most of the species belong to Rhinobatus. The best-known American species are Rhinobatus lentiginosus of Florida and Rhinobatus productus of California. The names guitar-fish, fiddler-fish, etc., refer to the form of the body. Numerous fossil species, allied to the recent forms, occur from the Jurassic. Species much like Rhinobatus occur in the Cretaceous and Eocene. Tamiobatis vetustus, lately described by Dr. Eastman from a skull found in the Devonian of eastern Kentucky, the oldest ray-like fish yet known, is doubtless the type of a distinct family, Tamiobatidæ. It is more likely a shark however than a ray, although the skull has a flattened ray-like form.
Rajidæ, or Skates.—The Rajidæ, skates, or rays, inhabit the colder waters of the globe and are represented by a large number of living species. In this family the tail is stout, with two-rayed dorsal fins and sometimes a caudal fin. The skin is variously armed with spines, there being always in the male two series of specialized spinous hooks on the outer edge of the pectoral fin. There is no serrated spine or "sting," and in all the species the eggs are laid in leathery cases, which are "wheelbarrow-shaped," with a projecting tube at each of the four angles. The size of this egg-case depends on the size of the species, ranging from three to about eight inches in length. In some species more than one egg is included in the same case.
Some of the species, known on our coasts as "barn-door skates," reach a length of four or five feet. Among these are Raja lævis and Raja ocellata on our Atlantic coast, Raja binoculata in California, and Raja tengu in Japan. The small tobacco-box skate, brown with black spots, abundant on the New England coast, is Raja erinacea. The corresponding species in California is Raja inornata, and in Japan Raja kenojei. Numerous other species, Raja batis, clavata, circularis, fullonica, etc., occur on the coasts of Europe. Some species are variegated in color, with eye-like spots or jet-black marblings. Still others, living in deep waters, are jet-black with the body very soft and limp. For these Garman has proposed the generic name Malacorhinus, a name which may come into general use when the species are better known. In the deep seas rays are found even under the equator. In the south-temperate zone the species are mostly generically distinct, Psammobatis being a typical form, differing from Raja. Discobatus sinensis, common in China and Japan, is a shagreen-covered form, looking like a Rhinobatus. It is, however, a true ray, laying its eggs in egg-cases, and with the pectorals extending on the snout. Fossil Rajidæ, known by the teeth and bony tubercles, are found from the Cretaceous onward. They belong to Raja and to the extinct genera Dynatobatis, Oncobatis, and Acanthobatis. The genus Arthropterus (rileyi) from the Lias, known from a large pectoral fin, with distinct cylindrical-jointed rays, may have been one of the Rajidæ, or perhaps the type of a distinct family, Arthropteridæ.
Narcobatidæ, or Torpedoes.—The torpedoes, or electric rays (Narcobatidæ), are characterized by the soft, perfectly smooth skin, by the stout tail with rayed fins, and by the ovoviviparous habit, the eggs being hatched internally. In all the species is developed an elaborate electric organ, muscular in its origin and composed of many hexagonal cells, each filled with soft fluid. These cells are arranged under the skin about the back of the head and at the base of the pectoral fin, and are capable of benumbing an enemy by means of a severe electric shock. The exercise of this power soon exhausts the animal, and a certain amount of rest is essential to recovery.
Petalodontidæ.—Near the Squatinidæ, between the sharks and the rays, Woodward places the large extinct family of Petalodontidæ, with coarsely paved teeth each of which is elongate with a central ridge and one or more strong roots at base. The best-known genera are Janassa and Petalodus, widely distributed in Carboniferous time. Janassa is a broad flat shark, or, perhaps, a skate, covered with smooth shagreen. The large pectoral fins are grown to the head; the rather large ventral fins are separated from them. The tail is small, and the fins, as in the rays, are without spines. The teeth bear some resemblance to those of Myliobatis. Janassa is found in the coal-measures of Europe and America, and other genera extend upward from the Subcarboniferous limestones, disappearing near the end of Carboniferous time. Petalodus is equally common, but known only from the teeth. Other widely distributed genera are Ctenoptychius and Polyrhizodus.
The common sting-rays, those in which the caudal spine is most developed, belong to the family of Dasyatidæ. This group is characterized by the small skate-like teeth and by the non-extension of the pectoral rays on the head. The skin is smooth or more or less rough. These animals lie flat on the sandy bottoms in nearly all seas, feeding on crabs and shellfish. All hatch the eggs within the body. The genus Urolophus has a rounded disk, and a stout, short tail with a caudal fin. It has a strong spine, and for its size is the most dangerous of the sting-rays. Urolophus halleri, the California species, was named for a young man who was stung by the species at the time of its first discovery at San Diego in 1863. Urolophus jamaicensis abounds in the West Indies, Urolophus mundus at Panama, and Urolophus fuscus in Japan. None of the species reach Europe. The true sting-ray (stingaree, or clam-cracker), Dasyatis, is more widely diffused and the species are very closely related. In these species the body is angular and the tail whip-like. Some of the species reach a length of ten or twelve feet. None have any economic value, and all are disliked by fishermen. Dasyatis pastinaca is common in Europe, Dasyatis centrura along our Atlantic coast, Dasyatis sabina ascends the rivers of Florida, and Dasyatis dipterura abounds in the bay of San Diego. Other species are found in tropical America, while still others (Dasyatis akajei, kuhlii, zugei, etc.) swarm in Japan and across India to Zanzibar.
Teeth and spines of all these genera are common as fossils from the Eocene onwards, as well as many of the extinct genus, Ptychodus, with cyclospondylous vertebræ. Ptychodus mammilaris, rugosus, and decurrens are characteristic of the Cretaceous of England. Myliobatis dixoni is common in the European Eocene, as is also Myliobatis toliapicus and Aëtobatis irregularis. Apocopodon seriacus is known from the Cretaceous of Brazil.
Family Psammodontidæ.—The Psammodontidæ are known only from the teeth, large, flat, or rounded and finely dotted or roughened on the upper surface, as the name Psammodus (ψάμμος, sand; ὀδούς, tooth) would indicate. The way in which the jaws lie indicates that these teeth belonged to rays rather than sharks. Numerous species have been described, mostly from the Subcarboniferous limestones. Archæobatis gigas, perhaps, as its name would indicate, the primeval skate, is from the Subcarboniferous limestone of Greencastle, Indiana. Teeth of numerous species of Psammodus and Copodus are found in many rocks of Carboniferous age. Psammodus rugosus common in Carboniferous rocks of Europe.
"The Holocephali have always been a doubtful group, anatomy and palæontology contributing but imperfect evidence as to their position in the gnathostome phylum. Their embryology, however, is still undescribed, except in a brief note by T. J. Parker, and it is reasonably looked to to contribute evidence as to their line of descent. The problem of the relationships of the Chimæroids has long been of especial interest to me, and it has led me to obtain embryonic material of a Pacific species of one of these forms. It may be of interest in this connection to state that the embryology of this form gives the clearest evidence that the wide separation of the Selachii and Holocephali is not tenable. The entire plan of development in Chimæra colliei is clearly like that of a shark. The ovulation is closely like that of certain of the rays and sharks: the eggs are large, the segmentation is distinctly shark-like; the circular blastoderm overgrows the yolk in an elasmobranchian manner. The early embryos are shark-like; and the later ones have, as T. J. Parker has shown, external gills, and I note further that these arise, precisely as in shark-embryos, from the posterior margin of the gill-bar. A spiracle also is present. A further and most interesting developmental feature is the fact that the autostylism in Chimæra is purely of secondary nature and is at the most of ordinal value. It is found that in a larva of Chimæra measuring 45 mm. in length, the palato-quadrate cartilage is still separated from the skull by a wide fissure. This becomes gradually reduced by the confluence of the palato-quadrate cartilage with the skull, the fusion taking place at both the anterior and posterior ends of the mesal rim of the cartilage. The remains of the fissure are still well marked in the young Chimæra, four inches in length; and a rudiment of it is present in the adult skull as a passage-way for a nerve. Regarding the dentition: it may also be noted in the present connection that the growth of the dental plates in Chimæra suggests distinctly elasmobranchian conditions. Thus on the roof of the mouth the palatine plates are early represented by a series of small more or less conical elements which resemble outwardly, at least, the 'anlagen' of the pavement teeth in cestraciont sharks."
Family Chimæridæ.—The existing Chimæras are known also as spookfishes, ratfishes, and elephant-fishes. These are divided by Garman into three families, and in the principal family, the Chimæridæ, the snout is blunt, the skin without plates, and the dorsal fin is provided with a long spine. The flat tritors vary in the different genera. The single genus represented among living fishes is Chimæra, found in cold seas and in the oceanic depths. The best-known species, Chimæra colliei, the elephant-fish, or chimæra of California, abounds in shallow waters of ten to twenty fathoms from Sitka to San Diego. It is a harmless fish, useless except for the oil in its liver, and of special interest to anatomists as the only member of the family to be found when desired for dissection. This species was first found at Monterey by Mr. Collie, naturalist of Captain Beechey's ship, the Blossom. It is brown in color, with whitish spots, and reaches a length of 2½ feet. As a shallow-water form, with certain differences in the claspers and in the tail, Chimæra colliei is sometimes placed in a distinct genus, Hydrolagus. Other species inhabit much greater depths and have the tail produced into a long filament. Of these, Chimæra monstrosa, the sea-cat of the north Atlantic, has been longer known than any other Chimæra. Chimæra affinis has been dredged in the Gulf Stream and off Portugal. Chimæra phantasma and Chimæra mitsukurii are frequently taken in Japan, and the huge jet-black Chimæra purpurascens in Hawaii and Japan. None of these species are valued as food, but all impress the spectator with their curious forms.
The best guess as to the affinities of the Ostracophores is perhaps that given by Dr. Ramsey H. Traquair ("Fossil Fishes of the Silurian Rocks of the South of Scotland," 1899). Traquair regards them as highly aberrant sharks, or, more exactly, as being derived, like the Chimæras, from a primitive Elasmobranch stock. In favor of this view is the character of their armature, the bony plates themselves to be regarded as formed by the fusion of shagreen grains or scales. According to Traquair: "Specialization from the most specialized form, Lanarkia, has been accompanied by (1) fusion of the spinelets (Lanarkia) or shagreen grains (Thelodus) into plates, scutes, and rhombic scales, supported by hard matter developed in a deeper layer of skin, and (2) alterations in the pectoral fin-flaps, which, becoming covered up by the postero-lateral plates in Drepanaspis, are finally no longer recognizable in the Pteraspidæ."
Order Heterostraci.—The Heterostraci (ἕτερος], different; ὀστράκος, box) have no bone-corpuscles in the coat of mail. This typically consists of a few pieces above, firmly united and traversed by dermal sense-organs or "lateral lines." The ventral shield is simple. Four families are recognized by Traquair as constituting the Heterostraci, these forming a continuous series from shark-like forms to the carapace-covered Pteraspis. In the most primitive family, the Thelodontidæ,[154] the head and trunk are covered with small scales or tubercles of dentine and not fused into large plates. The tail is slender and heterocercal, the caudal fin deeply forked. Until lately these tubercles were regarded as belonging to sharks, and they are still regarded by Traquair as evidence of the affinity of the Heterostraci with the Acanthodei. Dr. Traquair thinks that a flap or lappet-like projection behind the head may be a pectoral fin. The three known genera are Thelodus, Lanarkia, and Ateleaspis. In Thelodus the scales consist of a base and a crown separated by a constriction or neck. Thelodus scoticus, Thelodus pagei, and Thelodus planus are found in the Silurian rocks of Scotland. Other species, as Thelodus tulensis of Russia, extend to the Upper Devonian.
The family of Drepanaspidæ consists of a single recently known species, Drepanaspis gmundenensis, found in a pyritized condition in purple roofing-slate in Gmünden, Germany. This fish, which reaches a length of about two feet, has a broad head, with eyes on its outer margin, with a slender body and heterocercal tail. The head has a broad median plate and smaller polygonal ones. The flaps, supposed to represent the pectoral fins, are here cased in immovable bone. No trace of internal skeleton is found by Traquair, who has given the restoration of this species, but the mouth has been outlined.
Order Osteostraci.—The Osteostraci (ὄστεον, bone; οστρακος, box) (called Aspidocephali by Rohon) have bone-corpuscles in the shields, and the shield of the back is in one piece without lateral-line channels or sense-organs. Ventral shield single. The order includes three families. The Cephalaspidæ have the shields tuberculate, the one between the eyes fixed, and the anterior body-shields are not fused into a continuous plate. The best known of the numerous species is Cephalaspis lyelli from the Lower Devonian of England. Hemicyclaspis murchisoni occurs in the Upper Silurian of England, and the extraordinary Cephalaspis dawsoni in the Lower Devonian of Gaspé, Canada. Eukeraspis pustulifera has the head-shield very slender and armed with prickles. In the Thyestidæ the anterior body-scales are fused into a continuous plate. Thyestis and Didymaspis are genera of this type. The Odontotodontidæ (Tremataspidæ) have the shield truncate behind, its surface finely punctate, and the piece between the eyes not fixed. Odontotodus[155] schrenki is found in the Upper Silurian of the Island of Oesel in company with species of Thyestes. The Euphaneropidæ are represented in the Devonian of Quebec.
This group contains one family, the Asterolepidæ, with numerous species, mostly from Devonian rocks. The best known genus is Pterichthyodes,[156] in which the anterior median plate of the back is overlapped by the posterior dorso-lateral. Pterichthyodes milleri from the Lower Devonian, named by Agassiz for Hugh Miller, is the best known species, although numerous others, mostly from Scottish quarries, are in the British Museum. Asterolepis maximus is a very large species from the same region, known from a single plate. Bothriolepis canadensis is from the Upper Devonian of Scaumenac Bay near Quebec, numerous specimens and fragments finely preserved having been found.
The earliest remains of Ostracophori are found in Ordovician or Lower Silurian rocks of the Trenton horizon at Cañon City, Colorado. These consist of enormous numbers of small fragments of bones mixed with sand. With these is a portion of the head carapace of a small Ostracophore which has been named by Dr. Walcott Asteraspis desiderata and referred provisionally to the family of Asterolepidæ, which belongs otherwise to the Lower Devonian.
In the Ludlow shales we find the next appearance of the Ostracophores, two families, Thelodontidæ and Birkeniidæ, being there represented.
Order Anaspida.—Recently a fourth order, Anaspida (ἄ, without; ἀσπίς, shield), has been added to the Ostracophori through the researches of Dr. Traquair. This group occurs in the Upper Silurian in the south of Scotland. It includes the single family Birkeniidæ, characterized by the fusiform body, bluntly rounded head, bilobate, heterocercal tail, and a median row of hooked spinous scales along the ventral margin. No trace of jaws, teeth, limbs, or internal skeleton has been found. Unlike other Ostracophores, Birkenia has no cranial buckler with orbits on the top, nor have the scales and tubercles the microscopic structure found in other Ostracophores. In the genus Birkenia the head and body are completely covered by tubercular scutes. The gill-openings seem to be represented by a series of small perforations on the sides. A dorsal fin is present. Birkenia elegans is from the Ludlow and Downstonian rocks of southern Scotland. Lasianius problematicus from the same rocks is very similar, but is scaleless. It has a row of ventral plates like those of Birkenia, the only other hard parts it possesses being a number of parallel rods behind the head, homologous with the lateral series of Birkenia. Lasianius is therefore a specialized and degenerate representation of Birkenia, differing somewhat as "the nearly naked Phanerosteon differs from other Palæoniscidæ whose bodies are covered with osseous scales."
The head in all the species is covered with a great bony helmet. Behind this on the nape is another large shield, and between the two is usually a huge joint which Dr. Dean compares to the hinge of a spring-beetle (Elater).
"The puzzling characters of the Arthrodirans do not seem to be lessened by a more definite knowledge of their different forms. The tendency, as already noted, seems to be at present to regard the group provisionally as a widely modified offshoot of the primitive Dipnoans, basing this view upon their general structural characters, dermal plates, dentition, autostylism. But only in the latter regard could they have differed more widely from the primitive Elasmobranch or Teleostome, if it be admitted that in the matter of dermal structures they may be clearly separated from the Chimæroid. It certainly is difficult to believe that the articulation of the head of Arthrodirans could have been evolved after dermal bones had come to be formed, or that a Dipnoan could become so metamorphosed as to lose not only its body armoring, but its pectoral appendages as well. The size of the pectoral girdle is, of course, little proof that an anterior pair of fins must have existed, since this may well have been evolved in relation to the muscular supports of plastron, carapace, trunk, and head. The intermovement of the dental plates, seen especially in Dinichthys, is a further difficulty in accepting their direct descent from the Dipnoans."
Occurrence of Arthrodires.—These fishes occur in abundance from the Silurian times to the Mesozoic. In the Devonian their gigantic size and thick armor gave them the leading position among the hosts of the sea. Among the genera there occurred "series of forms most interesting as to their evolution." "It is found more and more evident," says Dr. Dean ("Fishes, Living and Fossil," pp. 135, 136) "that the Arthrodirans may have represented the dominant group in the Devonian period, as were the sharks in the Carboniferous, or as are the Teleosts in modern times. There were forms which, like Coccosteus, had eyes at the notches of the head-buckler; others, like Macropetalichthys, in which orbits were well centralized; some, like Dinichthys and Titanichthys, with the pineal foramen present; some with pectoral spines(?); some with elaborately sculptured dermal plates. Among their forms appear to have been those whose shape was apparently subcylindrical, adapted for swift swimming; others (Mylostoma) whose trunk was depressed to almost ray-like proportions. In size they varied from that of the perch to that of a basking shark. In dentition they presented the widest range in variation, from the formidable shear-like jaws of Dinichthys to the lip-like mandibles of Titanichthys, the tearing teeth of Trachosteus, the wonderfully forked tooth-bearing jaw-tips of Diplognathus, to the Cestraciont type, Mylostoma. The latter form has hitherto been known only from its dentition, but now proves to be, as Newberry and Smith Woodward suggested, a typical Arthrodiran."
The fossil types are very closely allied to the lung-fishes, and the two groups have no doubt a common origin in Silurian times. It is now usually considered that the Crossopterygian is more primitive than the lung-fish, though at the same time more nearly related to the Ganoids, and through them to the ordinary fishes.
In the true Amphibians the lungs are better developed than in the Crossopterygian or Dipnoan, although the lungs are finally lost in certain salamanders which breathe through epithelial cells. The gills lose, among the Amphibia, their primitive importance, although in Proteus anguineus of Austria and Necturus maculosus, the American "mud-puppy" or water-dog, these persist through life. The archipterygium, or primitive fin, gives place to the chiropterygium, or fingered arm. In this the basal segment of the archipterygium gives place to the humerus, the diverging segments seen in the most specialized type of archipterygium (Polypterus) become perhaps radius and ulna, the intermediate quadrate mass of cartilage possibly becoming carpal bones, and from these spring the joints called metacarpals and phalanges. In the Amphibians and all higher forms the shoulder-girdle retains its primitive insertion at a distance from the head, and the posterior limbs remain abdominal.
The Fins of Crossopterygians.—Dollo and Boulenger regard the heterocercal tail as a primitive form, the diphycercal form being a result of degradation, connected with its less extensive use as an organ of propulsion. Most writers who adopt the theory of Gegenbaur that the archipterygium is the primitive form of the pectoral fin are likely, however, to consider the diphycercal tail found associated with it in the Ichthyotomi, Dipneusti, Crossopterygii as the more primitive form of the tail. From this form the heterocercal tail of the higher sharks and Ganoids may be derived, this giving way in the process of development to the imperfectly homocercal tail of the salmon, the homocercal tail of the perch, and the isocercal tail of the codfish and its allies, the gephyrocercal and the leptocercal tail, tapering or whip-like, representing various stages of degeneration. Boulenger draws a distinction between the protocercal tail, the one primitively straight, and the diphycercal tail modified, like the homocercal tail, from an heterocercal ancestry.
The Osteolepidæ differ from the Megalichthyidæ mainly in the presence of enameled rhomboid scales, as in Polypterus and Lepisosteus. In Glyptopomus these scales are sculptured, in the others smooth. In Osteolepis, Thursius, Diplopterus, and Glyptopomus a pineal foramen is present on the top of the head. This is wanting in Parabatrachus (Megalichthys of authors). In Osteolepis, Thursius, and Parabatrachus the tail is heterocercal, while in Diplopterus and Glyptopomus it is diphycercal. Osteolepis macrolepidotus and numerous other species occur in the Lower Devonian. Diplopterus agassizii is common in the same horizon. Megalichthys hibberti is found in the coal-measures, and Glyptopomus minimus in the Upper Devonian. Palæosteus is another genus recently described.
Order Actinistia.—In the Actinistia there is a single fin-ray to each basal bone, the axonosts of each ray fused in a single piece. The notochord is persistent, causing the back-bone in fossils to appear hollow, the cartilaginous material leaving no trace in the rocks. The genera and species are numerous, ranging from the Subcarboniferous to the Upper Cretaceous, many of them belonging to Cœlacanthus, the chief genus of the single family Cœlacanthidæ. In Cœlacanthus the fin-rays are without denticles. Cœlacanthus granulatus is found in the European Permian. Cœlacanthus elegans of the coal-measures is found in America also. In Undina the anterior fin-rays are marked with tubercles. Undina penicillata and Undina gulo from the Triassic are well-preserved species. In Macropoma (lewesiensis) the fin-rays are robust, long, and little articulated. Other genera are Heptanema, Coccoderma, Libys, Diplurus, and Graphiurus. Diplurus longicaudatus was found by Newberry in the Triassic of New Jersey and Connecticut.
Order Cladistia.—In the Cladistia the axis of the pectoral limb is fan-shaped, made of two diversified bones joined by cartilage. The notochord is restricted and replaced by ossified vertebræ. The axonosts of the dorsal and anal are in regular series, each bearing a fin-ray. The order contains the single family Polypteridæ. In this group the pectoral fin is formed differently from that of the other Crossopterygians, being broad, its base of two diverging bones with cartilage between. This structure, more specialized than in any other of the Crossopterygians or Dipneusti, has been regarded by Gill and others, as above stated, as the origin of the fingered hand (chiropterygium) of the frogs and higher vertebrates. The base of the diverging bones has been identified as the antecedent of the humerus, the bones themselves as radius and ulna, while the intervening non-ossified cartilage breaks up into carpal bones, from which metacarpals and digits ultimately diverge. This hypothesis is open to considerable doubt. The nostrils, as in true fishes, are superior. The body in these fishes is covered with rhombic enameled scales, as in the garpike; the head is similarly mailed, but, in distinction from the garpike, the anterior rays of the dorsal are developed as isolated spines.
The external gill of the young, first discovered by Steindachner in 1869, consists of a fleshy axis bordered above and below by secondary branches, themselves fringed. In form and structure this resembles the external gills of amphibians. It is inserted, not on the gill-arches, but on the hyoid arch. Its origin is from the external skin. It can therefore not be compared morphologically with the gills of other fishes, nor with the pseudobranchiæ, but rather with the external gills of larval sharks. The vertebræ are very numerous and biconcave as in ordinary fishes. Each of the peculiar dorsal spines is primitively a single spine, not a finlet of several pieces, as some have suggested. The enameled, rhomboid scales are in movable oblique whorls, each scale interlocked with its neighbors.
In Dipterus there are no marginal teeth, and the tail is heterocercal, not diphycercal, as in the other Dipnoans generally. Numerous species of Dipterus occur in Devonian rocks. In these the jugular plate is present, as in Uronemus. Dipterus valenciennesi is the best-known European species. Dipterus nelsoni and numerous other species are found in the Chemung and other groups of Devonian rocks in America.
Family Ceratodontidæ.—The Ceratodontidæ have the teeth above and below developed as triangular plates, set obliquely each with several cusps on the outer margin. Nearly all the species, representing the genera Ceratodus, Gosfordia, and Conchopoma, are now extinct, the single genus Neoceratodus still existing in Australian rivers. Numerous fragments of Ceratodus are found in Mesozoic rocks in Europe, Colorado, and India, Ceratodus latissimus, figured by Agassiz in 1838, being the best-known species.
In 1870 the Barramunda of the rivers of Queensland was described by Krefft, who recognized its relationship to Ceratodus and gave it the name of Ceratodus forsteri. Later, generic differences were noticed, and it was separated as a distinct group by Castelnau in 1876, under the name of Neoceratodus (later called Epiceratodus by Teller). Neoceratodus forsteri and a second species, Neoceratodus miolepis, have been since very fully discussed by Dr. Günther and Dr. Krefft. They are known in Queensland as Barramunda. They inhabit the rivers known as Burnett, Dawson, and Mary, reaching a length of six feet, and being locally much valued as food. From the salmon-colored flesh, they are known to the settlers in Queensland as "salmon." According to Dr. Günther, "the Barramunda is said to be in the habit of going on land, or at least on mud-flats; and this assertion appears to be borne out by the fact that it is provided with a lung. However, it is much more probable that it rises now and then to the surface of the water in order to fill its lung with air, and then descends again until the air is so much deoxygenized as to render a renewal of it necessary. It is also said to make a grunting noise which may be heard at night for some distance. This noise is probably produced by the passage of the air through the œsophagus when it is expelled for the purpose of renewal. As the Barramunda has perfectly developed gills besides the lung, we can hardly doubt that, when it is in water of normal composition and sufficiently pure to yield the necessary supply of oxygen, these organs are sufficient for the purpose of breathing, and that the respiratory function rests with them alone. But when the fish is compelled to sojourn in thick muddy water charged with gases, which are the products of decomposing organic matter (and this must be the case very frequently during the droughts which annually exhaust the creeks of tropical Australia), it commences to breathe air with its lung in the way indicated above. If the medium in which it happens to be is perfectly unfit for breathing, the gills cease to have any function; if only in a less degree, the gills may still continue to assist in respiration. The Barramunda, in fact, can breathe by either gills or lung alone or by both simultaneously. It is not probable that it lives freely out of water, its limbs being much too flexible for supporting the heavy and unwieldy body and too feeble generally to be of much use in locomotion on land. However, it is quite possible that it is occasionally compelled to leave the water, although we cannot believe that it can exist without it in a lively condition for any length of time.
The next change of the embryo is strikingly amphibian-like; the medullary folds rise above the egg's surface, and, arching over, fuse their edges in the median dorsal line. The medullary folds are seen closely apposed in the median line; hindward, however, they are still separate, and through this opening the blastopore may yet be seen. At this stage primitive segments are shown; in the brain region the medullary folds are still slightly separated.
Lepidosirenidæ.—The family Lepidosirenidæ, representing the suborder Diplopneumona, is represented by two genera of mudfishes found in streams of Africa and South America. Lepidosiren paradoxa was discovered by Natterer in 1837 in tributaries of the Amazon. It was long of great rarity in collections, but quite recently large numbers have been obtained, and Dr. J. Graham Kerr of the University of Cambridge has given a very useful account of its structure and development. From his memoir we condense the following record of its habits as seen in the swamps in a region known as Gran Chaco, which lies under the Tropic of Capricorn. These swamps in the rainy season have a depth of from two to four feet, becoming entirely dry in the southern winter (June, July).
Kerr on the Habits of Lepidosiren.—The loalach, as the Lepidosiren is locally called, is normally sluggish, wriggling slowly about at the bottom of the swamp, using its hind limbs in irregular alternation as it clambers through the dense vegetation. More rapid movement is brought about by lateral strokes of the large and powerful posterior end of the body. It burrows with great facility, gliding through the mud, for which form of movement the shape of the head, with the upper lip overlapping the lower and the external nostril placed within the lower lip, is admirably adapted. It feeds on plants, algæ, and leaves of flower-plants. The gills are small and quite unable to supply its respiratory needs, and the animal must rise to the surface at intervals, like a frog. It breathes with its lungs as continuously and rhythmically as a mammal, the air being inhaled through the mouth. The animal makes no vocal sound, the older observation that it utters a cry like that of a cat being doubtless erroneous. Its strongest sense is that of smell. In darkness it grows paler in color, the black chromatophores shrinking in absence of light and enlarging in the sunshine. In injured animals this reaction becomes much less, as they remain pale even in daylight.
In the rainy season when food is abundant the Lepidosiren eats voraciously and stores great quantities of orange-colored fat in the tissues between the muscles. In the dry season it ceases to feed, or, as the Indians put it, it feeds on water. When the water disappears the Lepidosiren burrows down into the mud, closing its gill-openings, but breathing through the mouth. As the mud stiffens it retreats to the lower part of its burrow, where it lies with its tail folded over its face, the body surrounded by a mucous secretion. In its burrow there remains an opening which is closed by a lid of mud. At the end of the dry season this lid is pushed aside, and the animal comes out when the water is deep enough. When the waters rise the presence of Lepidosirens can be found only by a faint quivering movement of the grass in the bottom of the swamp. When taken the body is found to be as slippery as an eel and as muscular. The eggs are laid in underground burrows in the black peat. Their galleries run horizontally and are usually two feet long by eight inches wide. After the eggs are laid the male remains curled up in the nest with them. In the spawning season an elaborate brush is developed in connection with the ventral fins.
[Pg 1]
[Pg 2]

Fig. 1.—Long-eared Sunfish, Lepomis megalotis (Rafinesque). (From life by R. W. Shufeldt.)—Page 2.
CHAPTER I
THE LIFE OF THE FISH
A POPULAR ACCOUNT OF THE LIFE OF THE LONG-EARED SUNFISH, LEPOMIS MEGALOTIS

What is a Fish?—A fish is a back-boned animal which lives in the water and cannot ever live very long anywhere else. Its ancestors have always dwelt in water, and most likely its descendents will forever follow their example. So, as the water is a region very different from the fields or the woods, a fish in form and structure must be quite unlike all the beasts and birds that walk or creep or fly above ground, breathing air and being fitted to live in it. There are a great many kinds of animals called fishes, but in this all of them agree: all have some sort of a back-bone, all of them breathe their life long by means of gills, and none have fingers or toes with which to creep about on land.
The Long-eared Sunfish.—If we would understand a fish, we must first go and catch one. This is not very hard to do, for there are plenty of them in the little rushing brook or among the lilies of the pond. Let us take a small hook, put on it an angleworm or a grasshopper,—no need to seek an elaborate artificial fly,—and we will go out to the old "swimming-hole" or the deep eddy at the root of the old stump where the stream has gnawed away the bank in changing its course. Here we will find fishes, and one of them will take the bait very soon. In one part of the country the first fish that bites will be different from the first one taken in some other. But as we are fishing in the United States, we will locate our brook in the centre of population of our country. This will be to the northwest of Cincinnati, among the low wooded hills from which clear brooks flow over gravelly bottoms toward the Ohio River. Here we will catch sunfishes of certain species, or maybe rock bass or catfish: any of these will do for our purpose. But one of our sunfishes is especially beautiful—mottled blue and golden and scarlet, with a long, black, ear-like appendage backward from his gill-covers—and this one we will keep and hold for our first lesson in fishes. It is a small fish, not longer than your hand most likely, but it can take the bait as savagely as the best, swimming away with it with such force that you might think from the vigor of its pull that you have a pickerel or a bass. But when it comes out of the water you see a little, flapping, unhappy, living plate of brown and blue and orange, with fins wide-spread and eyes red with rage.

Fig. 2.—Long-eared Sunfish, Lepomis megalotis (Rafinesque). (From Clear Creek, Bloomington, Indiana.) Family Centrarchidæ.
Form of the Fish.—And now we have put the fish into a bucket of water, where it lies close to the bottom. Then we take it home and place it in an aquarium, and for the first time we have a chance to see what it is like. We see that its body is almost elliptical in outline, but with flat sides and shaped on the lower parts very much like a boat. This form we see is such as to enable it to part the water as it swims. We notice that its progress comes through the sculling motion of its broad, flat tail.
Face of a Fish.—When we look at the sunfish from the front we see that it has a sort of face, not unlike that of higher animals. The big eyes, one on each side, stand out without eyelids, but the fish can move them at will, so that once in a while he seems to wink. There isn't much of a nose between the eyes, but the mouth is very evident, and the fish opens and shuts it as it breathes. We soon see that it breathes water, taking it in through the mouth and letting it flow over the gills, and then out through the opening behind the gill-covers.
How the Fish Breathes.—If we take another fish—for we shall not kill this one—we shall see that in its throat, behind the mouth-cavity, there are four rib-like bones on each side, above the beginning of the gullet. These are the gill-arches, and on each one of them there is a pair of rows of red fringes called the gills. Into each of these fringes runs a blood-vessel. As the water passes over it the oxygen it contains is absorbed through the skin of the gill-fringe into the blood, which thus becomes purified. In the same manner the impurities of the blood pass out into the water, and go out through the gill-openings behind. The fish needs to breathe just as we do, though the apparatus of breathing is not the same. Just as the air becomes loaded with impurities when many people breathe it, so does the water in our jar or aquarium become foul if it is breathed over and over again by fishes. When a fish finds the water bad he comes to the surface to gulp air, but his gills are not well fitted to use undissolved air as a substitute for that contained in water. The rush of a stream through the air purifies the water, and so again does the growth of water plants, for these in the sunshine absorb and break up carbonic acid gas, and throw out oxygen into the water.
Teeth of the Fish.—On the inner side of the gill-arch we find some little projections which serve as strainers to the water. These are called gill-rakers. In our sunfish they are short and thick, seeming not to amount to much but in a herring they are very long and numerous.
Behind the gills, at the opening of the gullet, are some roundish bones armed with short, thick teeth. These are called pharyngeals. They form a sort of jaws in the throat, and they are useful in helping the little fish to crack shells. If we look at the mouth of our live fish, we shall find that when it breathes or bites it moves the lower jaw very much as a dog does. But it can move the upper jaw, too, a little, and that by pushing it out in a queer fashion, as though it were thrust out of a sheath and then drawn in. If we look at our dead fish, we shall see that the upper jaw divides in the middle and has two bones on each side. On one bone are rows of little teeth, while the other bone that lies behind it has no teeth at all. The lower jaw has little teeth like those of the upper jaw, and there is a patch of teeth on the roof of the mouth also. In some sunfishes there are three little patches, the vomer in the middle and the palatines on either side.
The tongue of the fish is flat and gristly. It cannot move it, scarce even taste its food with it, nor can it use it for making a noise. The unruly member of a fish is not its tongue, but its tail.
How the Fish Sees.—To come back to the fish's eye again. We say that it has no eyelids, and so, if it ever goes to sleep, it must keep its eyes wide open. The iris is brown or red. The pupil is round, and if we could cut open the eye we should see that the crystalline lens is almost a perfect sphere, much more convex than the lens in land animals. We shall learn that this is necessary for the fish to see under water. It takes a very convex lens or even one perfectly round to form images from rays of light passing through the water, because the lens is but little more dense than the water itself. This makes the fish near-sighted. He cannot see clearly anything out of water or at a distance. Thus he has learned that when, in water or out, he sees anything moving quickly it is probably something dangerous, and the thing for him to do is to swim away and hide as swiftly as possible.
In front of the eye are the nostrils, on each side a pair of openings. But they lead not into tubes, but into a little cup lined with delicate pink tissues and the branching nerves of smell. The organ of smell in nearly all fishes is a closed sac, and the fish does not use the nostrils at all in breathing. But they can indicate the presence of anything in the water which is good to eat, and eating is about the only thing a fish cares for.
Color of the Fish.—Behind the eye there are several bones on the side of the head which are more or less distinct from the skull itself. These are called membrane bones because they are formed of membrane which has become bony by the deposition in it of salts of lime. One of these is called the opercle, or gill-cover, and before it, forming a right angle, is the preopercle, or false gill-cover. On our sunfish we see that the opercle ends behind in a long and narrow flap, which looks like an ear. This is black in color, with an edging of scarlet as though a drop of blood had spread along its margin. When the fish is in the water its back is dark greenish-looking, like the weeds and the sticks in the bottom, so that we cannot see it very plainly. This is the way the fish looks to the fishhawks or herons in the air above it who may come to the stream to look for fish. Those fishes which from above look most like the bottom can most readily hide and save themselves. The under side of the sunfish is paler, and most fishes have the belly white. Fishes with white bellies swim high in the water, and the fishes who would catch them lie below. To the fish in the water all outside the water looks white, and so the white-bellied fishes are hard for other fishes to see, just as it is hard for us to see a white rabbit bounding over the snow.

Fig. 3.—Common sunfish, Eupomotis gibbosus (Linnæus). Natural size. (From life by R. W. Shufeldt.)
But to be known of his own kind is good for the sunfish, and we may imagine that the black ear-flap with its scarlet edge helps his mate and friends to find him out, where they swim on his own level near the bottom. Such marks are called recognition-marks, and a great many fishes have them, but we have no certain knowledge as to their actual purpose.
We are sure that the ear-flap is not an ear, however. No fishes have any external ear, all their hearing apparatus being buried in the skull. They cannot hear very much: possibly a great jar or splash in the water may reach them, but whenever they hear any noise they swim off to a hiding-place, for any disturbance whatever in the water must arouse a fish's anxiety. The color of the live sunfish is very brilliant. Its body is covered with scales, hard and firm, making a close coat of mail, overlapping one another like shingles on a roof. Over these is a thin skin in which are set little globules of bright-colored matter, green, brown, and black, with dashes of scarlet, blue, and white as well. These give the fish its varied colors. Some coloring matter is under the scales also, and this especially makes the back darker than the lower parts. The bright colors of the sunfish change with its surroundings or with its feelings. When it lies in wait under a dark log its colors are very dark. When it rests above the white sands it is very pale. When it is guarding its nest from some meddling perch its red shades flash out as it stands with fins spread, as though a water knight with lance at rest, looking its fiercest at the intruder.
When the sunfish is taken out of the water its colors seem to fade. In the aquarium it is generally paler, but it will sometimes brighten up when another of its own species is placed beside it. A cause of this may lie in the nervous control of the muscles at the base of the scales. When the scales lie very flat the color has one appearance. When they rise a little the shade of color seems to change. If you let fall some ink-drops between two panes of glass, then spread them apart or press them together, you will see changes in the color and size of the spots. Of this nature is the apparent change in the colors of fishes under different conditions. Where the fish feels at its best the colors are the richest. There are some fishes, too, in which the male grows very brilliant in the breeding season through the deposition of red, white, black, or blue pigments, or coloring matter, on its scales or on its head or fins, this pigment being absorbed when the mating season is over. This is not true of the sunfish, who remains just about the same at all seasons. The male and female are colored alike and are not to be distinguished without dissection. If we examine the scales, we shall find that these are marked with fine lines and concentric striæ, and part of the apparent color is due to the effect of the fine lines on the light. This gives the bluish lustre or sheen which we can see in certain lights, although we shall find no real blue pigment under it. The inner edge of each scale is usually scalloped or crinkled, and the outer margin of most of them has little prickly points which make the fish seem rough when we pass our hand along his sides.

Fig. 4.—Ozorthe dictyogramma (Herzenstein). A Japanese blenny, from Hakodate: showing increased number of lateral lines, a trait characteristic of many fishes of the north Pacific.
The Lateral Line.—Along the side of the fish is a line of peculiar scales which runs from the head to the tail. This is called the lateral line. If we examine it carefully, we shall see that each scale has a tube from which exudes a watery or mucous fluid. Behind these tubes are nerves, and although not much is known of the function of the tubes, we can be sure that in some degree the lateral line is a sense-organ, perhaps aiding the fish to feel sound-waves or other disturbances in the water.
The Fins of the Fish.—The fish moves itself and directs its course in the water by means of its fins. These are made up of stiff or flexible rods growing out from the body and joined together by membrane. There are two kinds of these rays or rods in the fins. One sort is without joints or branches, tapering to a sharp point. The rays thus fashioned are called spines, and they are in the sunfish stiff and sharp-pointed. The others, known as soft rays, are made up of many little joints, and most of them branch and spread out brush-like at their tips. In the fin on the back the first ten of the rays are spines, the rest are soft rays. In the fin under the tail there are three spines, and in each fin at the breast there is one spine with five soft rays. In the other fins all the rays are soft.
The fin on the back is called the dorsal fin, the fin at the end of the tail is the caudal fin, the fin just in front of this on the lower side is the anal fin. The fins, one on each side, just behind the gill-openings are called the pectoral fins. These correspond to the arms of man, the wings of birds, or the fore legs of a turtle or lizard. Below these, corresponding to the hind legs, is the pair of fins known as the ventral fins. If we examine the bones behind the gill-openings to which the pectoral fins are attached, we shall find that they correspond after a fashion to the shoulder-girdle of higher animals. But the shoulder-bone in the sunfish is joined to the back part of the skull, so that the fish has not any neck at all. In animals with necks the bones at the shoulder are placed at some distance behind the skull.
If we examine the legs of a fish, the ventral fins, we shall find that, as in man, these are fastened to a bone inside called the pelvis. But the pelvis in the sunfish is small and it is placed far forward, so that it is joined to the tip of the "collar-bone" of the shoulder-girdle and pelvis attached together. The caudal fin gives most of the motion of a fish. The other fins are mostly used in maintaining equilibrium and direction. The pectoral fins are almost constantly in motion, and they may sometimes help in breathing by starting currents outside which draw water over the gills.
The Skeleton of the Fish.—The skeleton of the fish, like that of man, is made up of the skull, the back-bone, the limbs, and their appendages. But in the fish the bones are relatively smaller, more numerous, and not so firm. The front end of the vertebral column is modified as a skull to contain the little brain which serves for all a fish's activities. To the skull are attached the jaws, the membrane bones, and the shoulder-girdle. The back-bone itself in the sunfish is made of about twenty-four pieces, or vertebræ. Each of these has a rounded central part, concave in front and behind. Above this is a channel through which the great spinal cord passes, and above and below are a certain number of processes or projecting points. To some of these, through the medium of another set of sharp bones, the fins of the back are attached. Along the sides of the body are the slender ribs.
The Fish in Action.—The fish is, like any other animal, a machine to convert food into power. It devours other animals or plants, assimilates their substance, takes it over into itself, and through its movements uses up this substance again. The food of the sunfish is made up of worms, insects, and little fishes. To seize these it uses its mouth and teeth. To digest them it needs its alimentary canal, made of the stomach with its glands and intestines. If we cut the fish open, we shall find the stomach with its pyloric cæca, near it the large liver with its gall-bladder, and on the other side the smaller spleen. After the food is dissolved in the stomach and intestines the nutritious part is taken up by the walls of the alimentary canal, whence it passes into the blood.
The blood is made pure in the gills, as we have already seen. To send it to the gills the fish has need of a little pumping-engine, and this we shall find at work in the fish as in all higher animals. This engine of stout muscle surrounding a cavity is called the heart. In most fishes it is close behind the gills. It contains one auricle and one ventricle only, not two of each as in man. The auricle receives the impure blood from all parts of the body. It passes it on to the ventricle, which, being thick-walled, is dark red in color. This passes the blood by convulsive action, or heart-beating, on to the gills. From these the blood is collected in arteries, and without again returning to the heart it flows all through the body. The blood in the fish flows sluggishly. The combustion of waste material goes on slowly, and so the blood is not made hot as it is in the higher beasts and birds. Fishes have relatively little blood; what there is is rather pale and cold and has no swift current.
If we look about in the inside of a fish, we shall find close along the lower side of the back-bone, covering the great artery, the dark red kidneys. These strain out from the blood a certain class of impurities, poisons made from nerve or muscle waste which cannot be burned away by the oxygen of respiration.
The Air-bladder.—In the front part of the sunfish, just above the stomach, is a closed sac, filled with air. This is called the air-bladder, or swim-bladder. It helps the fish to maintain its place in the water. In bottom fishes it is almost always small, while fishes that rise and fall in the current generally have a large swim-bladder. The gas inside it is secreted from the blood, for the sunfish has no way of getting any air into it from the outside.
But the primal purpose of the air-bladder was not to serve as a float. In very old-fashioned fishes it has a tube connecting it with the throat, and instead of being an empty sac it is a true lung made up of many lobes and parts and lined with little blood-vessels. Such fishes as the garpike and the bowfin have lung-like air-bladders and gulp air from the surface of the water.
In the very little sunfish, when he is just hatched, the air-bladder has an air-duct, which, however, is soon lost, leaving only a closed sac. From all this we know that the air-bladder is the remains of what was once a lung, or additional arrangement for breathing. As the gills furnish oxygen enough, the lung of the common fish has fallen into disuse and thrifty Nature has used the parts and the space for another and a very different purpose. This will serve to help us to understand the swim-bladder and the way the fish came to acquire it as a substitute for a lung.
The Brain of the Fish.—The movements of the fish, like those of every other complex animal, are directed by a central nervous system, of which the principal part is in the head and is known as the brain. From the eye of the fish a large nerve goes to the brain to report what is in sight. Other nerves go from the nostrils, the ears, the skin, and every part which has any sort of capacity for feeling. These nerves carry their messages inward, and when they reach the brain they may be transformed into movement. The brain sends back messages to the muscles, directing them to contract. Their contraction moves the fins, and the fish is shoved along through the water. To scare the fish or to attract it to its food or to its mate is about the whole range of the effect that sight or touch has on the animal. These sensations changed into movement constitute what is called reflex action, performance without thinking of what is being done. With a boy, many familiar actions may be equally reflex. The boy can also do many other things "of his own accord," that is, by conscious effort. He can choose among a great many possible actions. But a fish cannot. If he is scared, he must swim away, and he has no way to stop himself. If he is hungry, and most fishes are so all the time, he will spring at the bait. If he is thirsty, he will gasp, and there is nothing else for him to do. In other words, the activities of a fish are nearly all reflex, most of them being suggested and immediately directed by the influence of external things. Because its actions are all reflex the brain is very small, very primitive, and very simple, nothing more being needed for automatic movement. Small as the fish's skull-cavity is, the brain does not half fill it.

Fig. 5.—Common Sunfish, Eupomotis gibbosus (Linnæus). Natural size. (From life by R. W. Shufeldt.)—Page 13.
The vacant space about the little brain is filled with a fatty fluid mass looking like white of egg, intended for its protection. Taking the dead sunfish (for the live one we shall look after carefully, giving him every day fresh water and a fresh worm or snail or bit of beef), if we cut off the upper part of the skull we shall see the separate parts of the brain, most of them lying in pairs, side by side, in the bottom of the brain-cavity. The largest pair is near the middle of the length of the brain, two nerve-masses (or ganglia), each one round and hollow. If we turn these over, we shall see that the nerves of the eye run into them. We know then that these nerve-masses receive the impressions of sight, and so they are called optic lobes. In front of the optic lobes are two smaller and more oblong nerve-masses. These constitute the cerebrum. This is the thinking part of the brain, and in man and in the higher animals it makes up the greater part of it, overlapping and hiding the other ganglia. But the fish has not much need for thinking and its fore-brain or cerebrum is very small. In front of these are two small, slim projections, one going to each nostril. These are the olfactory lobes which receive the sensation of smell. Behind the optic lobes is a single small lobe, not divided into two. This is the cerebellum and it has charge of certain powers of motion. Under the cerebellum is the medulla, below which the spinal cord begins. The rest of the spinal cord is threaded through the different vertebræ back to the tail, and at each joint it sends out nerves of motion and receives nerves of sense. Everything that is done by the fish, inside or outside, receives the attention of the little branches of the great nerve-cord.
The Fish's Nest.—The sunfish in the spawning time will build some sort of a nest of stones on the bottom of the eddy, and then, when the eggs are laid, the male with flashing eye and fins all spread will defend the place with a good deal of spirit. All this we call instinct. He fights as well the first time as the last. The pressure of the eggs suggests nest-building to the female. The presence of the eggs tells the male to defend them. But the facts of the nest-building and nest protection are not very well understood, and any boy who can watch them and describe them truly will be able to add something to science.
CHAPTER II
THE EXTERIOR OF THE FISH

Form of Body.—With a glance at the fish as a living organism and some knowledge of those structures which are to be readily seen without dissection, we are prepared to examine its anatomy in detail, and to note some of the variations which may be seen in different parts of the great group.
In general fishes are boat-shaped, adapted for swift progress through the water. They are longer than broad or deep and the greatest width is in front of the middle, leaving the compressed paddle-like tail as the chief organ of locomotion.
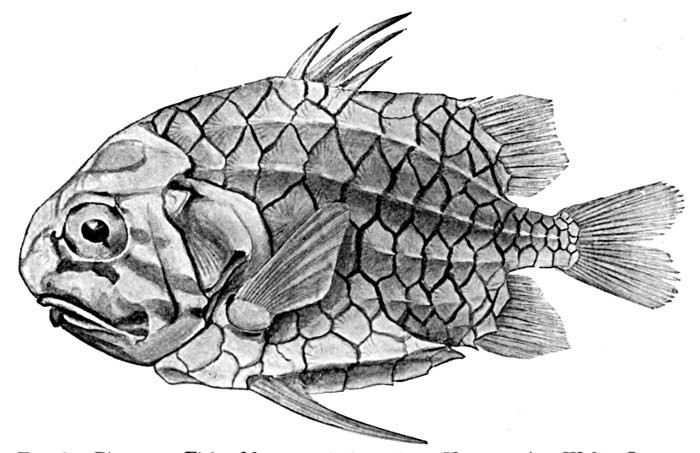
Fig. 6.—Pine-cone Fish, Monocentris japonicus (Houttuyn). Waka, Japan.
But to all these statements there are numerous exceptions. Some fishes depend for protection, not on swiftness, but on the thorny skin or a bony coat of mail. Some of these are almost globular in form, and their outline bears no resemblance to that of a boat. The trunkfish (Ostracion) in a hard bony box has no need of rapid progress.
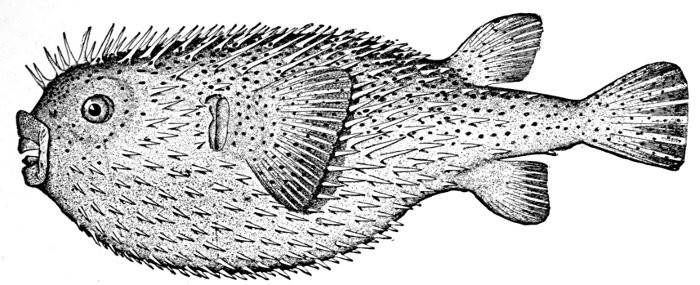
Fig. 7.—Porcupine-fish, Diodon hystrix (Linnæus). Tortugas Islands.

Fig. 8.—Thread-eel, Nemichthys avocetta Jordan and Gilbert. Vancouver Island.
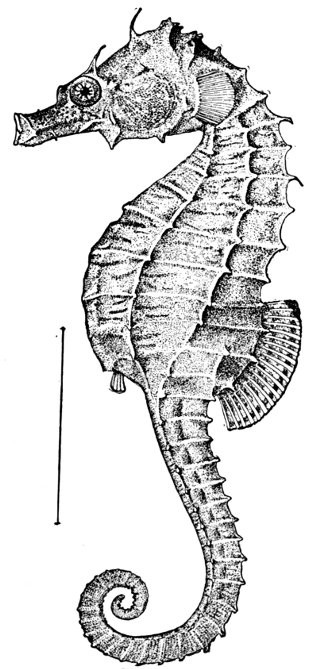
Fig. 9.—Sea-horse, Hippocampus hudsonius Dekay. Virginia.
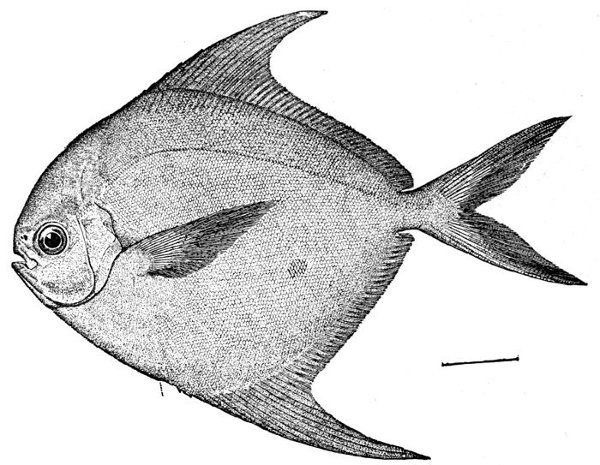
Fig. 10.—Harvest-fish, Peprilus paru (Linnæus). Virginia.
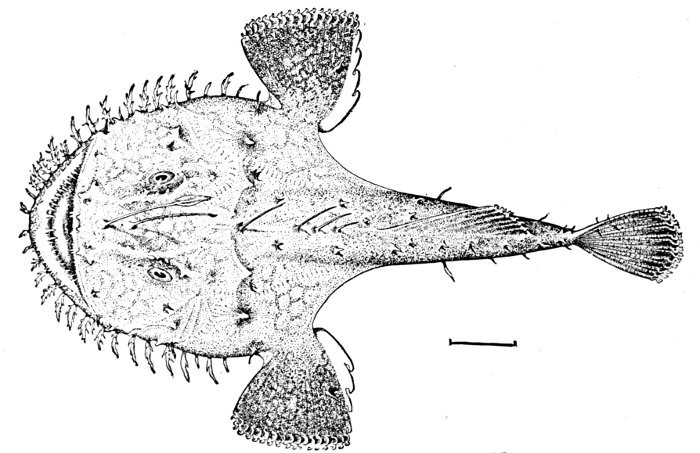
Fig. 11.—Anko or Fishing-frog, Lophius litulon (Jordan). Matsushima Bay, Japan. (The short line in all cases shows the degree of reduction; it represents an inch of the fish's length.)
The pine-cone fish (Monocentris japonicus) adds strong fin-spines to its bony box, and the porcupine fish (Diodon hystrix) is covered with long prickles which keep away all enemies.
Among swift fishes, there are some in which the body is much deeper than long, as in Antigonia. Certain sluggish fishes seem to be all head and tail, looking as though the body by some accident had been omitted. These, like the headfish (Mola mola) are protected by a leathery skin. Other fishes, as the eels, are extremely long and slender, and some carry this elongation to great extremes. Usually the head is in a line with the axis of the body, but in some cases, as the sea-horse (Hippocampus), the head is placed at right angles to the axis, and the body itself is curved and cannot be straightened without injury. The type of the swiftest fish is seen among the mackerels and tunnies, where every outline is such that a racing yacht might copy it.
The body or head of the fish is said to be compressed when it is flattened sidewise, depressed when it is flattened vertically. Thus the Peprilus (Fig. 10) is said to be compressed, while the fishing-frog (Lophius) (Fig. 11) has a depressed body and head. Other terms as truncate (cut off short), attenuate (long-drawn out), robust, cuboid, filiform, and the like may be needed in descriptions.
Measurement of the Fish.—As most fishes grow as long as they live, the actual length of a specimen has not much value for purposes of description. The essential point is not actual length, but relative length. The usual standard of measurement is the length from the tip of the snout to the base of the caudal fin. With this length the greatest depth of the body, the greatest length of the head, and the length of individual parts may be compared. Thus in the Rock Hind (Epinephelus adscensionis), fig. 12, the head is contained 2-3/5 times in the length, while the greatest depth is contained three times.
Thus, again, the length of the muzzle, the diameter of the eye, and other dimensions may be compared with the length of the head. In the Rock Hind, fig. 12, the eye is 5 in head, the snout is 4-2/5 in head, and the maxillary 2-3/5. Young fishes have the eye larger, the body slenderer, and the head larger in proportion than old fishes of the same kind. The mouth grows larger with age, and is sometimes larger also in the male sex. The development of the fins often varies a good deal in some fishes with age, old fishes and male fishes having higher fins when such differences exist. These variations are soon understood by the student of fishes and cause little doubt or confusion in the study of fishes.

Fig. 12.—Rock Hind or Cabra Mora of the West Indies, Epinephelus adscensionis (Osbeck). Family Serranidæ.
The Scales, or Exoskeleton.—The surface of the fish may be naked as in the catfish, or it may be covered with scales, prickles, shagreen, or bony plates. The hard covering of the skin, when present, is known as the exoskeleton, or outer skeleton. In the fish, the exoskeleton, whatever form it may assume, may be held to consist of modified scales, and this is usually obviously the case. The skin of the fish may be thick or thin, bony, horny, leathery, or papery, or it may have almost any intermediate character. When protected by scales the skin is usually thin and tender; when unprotected it may be ossified, as in the sea-horse; horny, as in the headfish; leathery, as in the catfish; or it may, as in the sea-snails, form a loose scarf readily detachable from the muscles below.
The scales themselves may be broadly classified as ctenoid, cycloid, placoid, ganoid, or prickly.
Ctenoid and Cycloid Scales.—Normally formed scales are rounded in outline, marked by fine concentric rings, and crossed on the inner side by a few strong radiating ridges and folds. They usually cover the body more or less evenly and are imbricated like shingles on a roof, the free edge being turned backward. Such normal scales are of two types, ctenoid or cycloid. Ctenoid scales have a comb-edge of fine prickles or cilia; cycloid scales have the edges smooth. These two types are not very different, and the one readily passes into the other, both being sometimes seen on different parts of the same fish. In general, however, the more primitive representatives of the typical fishes, those with abdominal ventrals and without spines in the fins, have cycloid or smooth scales. Examples are the salmon, herring, minnow, and carp. Some of the more specialized spiny-rayed fishes, as the parrot-fishes, have, however, scales equally smooth, although somewhat different in structure. Sometimes, as in the eel, the cycloid scales may be reduced to mere rudiments buried in the skin.
Ctenoid scales are beset on the free edge by little prickles or points, sometimes rising to the rank of spines, at other times soft and scarcely noticeable, when they are known as ciliate or eyelash-like. Such scales are possessed in general by the more specialized types of bony fishes, as the perch and bass, those with thoracic ventrals and spines in the fins.

Fig. 13.—Scales of Acanthoessus bronni (Agassiz). (After Dean.)
Placoid Scales.—Placoid scales are ossified papillæ, minute, enamelled, and close-set, forming a fine shagreen. These are characteristic of the sharks; and in the most primitive sharks the teeth are evidently modifications of these primitive structures. Some other fishes have scales which appear shagreen-like to sight and feeling, but only the sharks have the peculiar structure to which Agassiz gave the name of placoid. The rough prickles of the filefishes and some sculpins are not placoid, but are reduced or modified ctenoid scales, scales narrowed and reduced to prickles.
Bony and Prickly Scales.—Bony and prickly scales are found in great variety, and scarcely admit of description or classification. In general, prickly points on the skin are modifications of ctenoid scales. Ganoid scales are thickened and covered with bony enamel, much like that seen in teeth, otherwise essentially like cycloid scales. These are found in the garpike and in many genera of extinct Ganoid and Crossopterygian fishes. In the line of descent the placoid scale preceded the ganoid, which in turn was followed by the cycloid and lastly by the ctenoid scale. Bony scales in other types of fishes may have nothing structurally in common with ganoid scales or plates, however great may be the superficial resemblance.
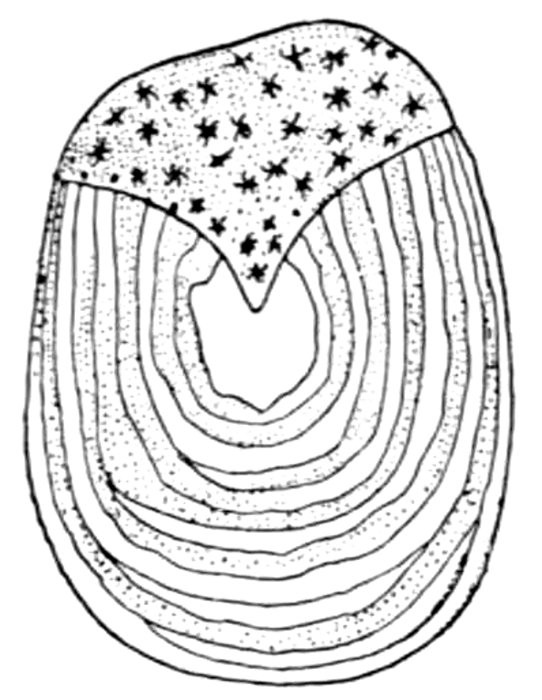
Fig. 14.—Cycloid Scale.
The distribution of scales on the body may vary exceedingly. In some fishes the scales are arranged in very regular series; in others they are variously scattered over the body. Some are scaly everywhere on head, body, and fins. Others may have only a few lines or patches. The scales may be everywhere alike, or they may in one part or another be greatly modified. Sometimes they are transformed into feelers or tactile organs. The number of scales is always one of the most valuable of the characters by which to distinguish species.
Lateral Line.—The lateral line in most fishes consists of a series of modified scales, each one provided with a mucous tube extending along the side of the body from the head to the caudal fin. The canal which pierces each scale is simple at its base, but its free edge is often branched or ramified. In most spiny-rayed fishes it runs parallel with the outline of the back. In most soft-rayed fishes it follows rather the outline of the belly. It is subject to many variations. In some large groups (Gobiidæ, Pæciliidæ) its surface structures are entirely wanting. In scaleless fishes the mucous tube lies in the skin itself. In some groups the lateral line has a peculiar position, as in the flying-fishes, where it forms a raised ridge bounding the belly. In many cases the lateral line has branches of one sort or another. It is often double or triple, and in some cases the whole back and sides of the fish are covered with lateral lines and their ramifications. Sometimes peculiar sense-organs and occasionally eye-like luminous spots are developed in connection with the lateral line, enabling the fish to see in the black depths of the sea. These will be noticed in another chapter.
The Lateral Line as a Mucous Channel.—The more primitive condition of the lateral line is seen in the sharks and chimæras, in which fishes it appears as a series of channels in or under the skin. These channels are filled with mucus, which exudes through occasional open pores. In many fishes the bones of the skull are cavernous, that is, provided with cavities filled with mucus. Analogous to these cavities are the mucous channels which in primitive fishes constitute the lateral line.
Fig. 15.—Singing Fish (with many lateral lines), Porichthys porosissimus (Cuv. and Val.). Gulf of Mexico.
Function of the Lateral Line.—The general function of the lateral line with its tubes and pores is still little understood. As the structures of the lateral line are well provided with nerves, it has been thought to be an organ of sense of some sort not yet understood. Its close relation to the ear is beyond question, the ear-sac being an outgrowth from it.
"The original significance of the lateral line," according to Dr. Dean,[2] "as yet remains undetermined. It appears intimately if not genetically related to the sense-organs of the head and gill region of the ancestral fish. In response to special aquatic needs, it may thence have extended farther and farther backward along the median line of the trunk, and in its later differentiation acquired its metameral characters." In view of its peculiar nerve-supply, "the precise function of this entire system of organs becomes especially difficult to determine. Feeling, in its broadest sense, has safely been admitted as its possible use. Its close genetic relationship to the hearing organ suggests the kindred function of determining waves of vibration. These are transmitted in so favorable a way in the aquatic medium that from the side of theory a system of hypersensitive end-organs may well have been established. The sensory tracts along the sides of the body are certainly well situated to determine the direction of the approach of friend, enemy, or prey."
The Fins of Fishes.—The organs of locomotion in the fishes are knows as fins. These are composed of bony or cartilaginous rods or rays connected by membranes. The fins are divided into two groups, paired fins and vertical fins. The pectoral fins, one on either side, correspond to the anterior limbs of the higher vertebrates. The ventral fins below or behind them represent the hinder limbs. Either or both pairs may be absent, but the ventrals are much more frequently abortive than the pectorals. The insertion of the ventral fins may be abdominal, as in the sharks and the more generalized of the bony fishes, thoracic under the breast (the pelvis attached to the shoulder-girdle) or jugular, under the throat. When the ventral fins are abdominal, the pectoral fins are usually placed very low. The paired fins are not in general used for progression in the water, but serve rather to enable the fish to keep its equilibrium. With the rays, however, the wing-like pectoral fins form the chief organ of locomotion.
The fin on the median line of the back is called the dorsal, that on the tail the caudal, and that on the lower median line the anal fin. The dorsal is often divided into two fins or even three. The anal is sometimes divided, and either dorsal or anal fin may have behind it detached single rays called finlets.
The rays composing the fin may be either simple or branched. The branched rays are always articulated, that is, crossed by numerous fine joints which render them flexible. Simple rays are also sometimes articulate. Rays thus jointed are known as soft rays, while those rays which are neither jointed nor branched are called spines. A spine is usually stiff and sharp-pointed, but it may be neither, and some spines are very slender and flexible, the lack of branches or joints being the feature which distinguishes spine from soft ray.
The anterior rays of the dorsal and anal fins are spinous in most fishes with thoracic ventrals. The dorsal fin has usually about ten spines, the anal three, but as to this there is much variation in different groups. When the dorsal is divided all the rays of the first dorsal and usually the first ray of the second are spines. The caudal fin has never true spines, though at the base of its lobes are often rudimentary rays which resemble spines. Most spineless fishes have such rudiments in front of their vertical fins. The pectoral, as a rule, is without spines, although in the catfishes and some others a single large spine may be developed. The ventrals when abdominal are usually without spines. When thoracic each usually, but not always, consists of one spine and five soft rays. When jugular the number of soft rays may be reduced, this being a phase of degeneration of the fin. In writing descriptions of fishes the number of spines may be indicated by Roman numerals, those of the soft rays by Arabic. Thus D. XII-I, 17 means that the dorsal is divided, that the anterior portion consists of twelve spines, the posterior of one spine and seventeen soft rays. In some fishes, as the catfish or the salmon, there is a small fin on the back behind the dorsal fin. This is known as the adipose fin, being formed of fatty substance covered by skin. In a few catfishes, this adipose fin develops a spine or soft rays.
Muscles.—The movements of the fins are accomplished by the muscles. These organs lie along the sides of the body, forming the flesh of the fish. They are little specialized, and not clearly differentiated as in the higher vertebrates.
With the higher fishes there are several distinct systems of muscles controlling the jaws, the gills, the eye, the different fins, and the body itself. The largest of all is the great lateral muscle, composed of flake-like segments (myocommas) which correspond in general with the number of the vertebræ. In general the muscles of the fish are white in color. In some groups, especially of the mackerel family, they are deep red, charged with animal oils. In the salmon they are orange-red, a color also due to the presence of certain oils.
In a few fishes muscular structures are modified into electric organs. These will be discussed in a later chapter.
FOOTNOTES:
[2] Fishes Recent and Fossil, p. 52.
CHAPTER III
THE DISSECTION OF THE FISH
[2] Fishes Recent and Fossil, p. 52.
"The original significance of the lateral line," according to Dr. Dean,[2] "as yet remains undetermined. It appears intimately if not genetically related to the sense-organs of the head and gill region of the ancestral fish. In response to special aquatic needs, it may thence have extended farther and farther backward along the median line of the trunk, and in its later differentiation acquired its metameral characters." In view of its peculiar nerve-supply, "the precise function of this entire system of organs becomes especially difficult to determine. Feeling, in its broadest sense, has safely been admitted as its possible use. Its close genetic relationship to the hearing organ suggests the kindred function of determining waves of vibration. These are transmitted in so favorable a way in the aquatic medium that from the side of theory a system of hypersensitive end-organs may well have been established. The sensory tracts along the sides of the body are certainly well situated to determine the direction of the approach of friend, enemy, or prey."

The Blue-green Sunfish.—The organs found in the abdominal cavity of the fish may be readily traced in a rapid dissection. Any of the bony fishes may be chosen, but for our purposes the sunfish will serve as well as any. The names and location of the principal organs are shown in the accompanying figure, from Kellogg's Zoology. It represents the blue-green sunfish, Apomotis cyanellus, from the Kansas River, but in these regards all the species of sunfishes are alike. We may first glance at the different organs as shown in the sequence of dissection, leaving a detailed account of each to the subsequent pages.
The Viscera.—Opening the body cavity of the fish, as shown in the plate, we see below the back-bone a membranous sac closed and filled with air. This is the air-bladder, a rudiment of that structure which in higher vertebrates is developed as a lung. The alimentary canal passes through the abdominal cavity extending from the mouth through the pharynx and ending at the anus or vent. The stomach has the form of a blind sac, and at its termination are a number of tubular sacs, the pyloric cæca, which secrete a digestive fluid. Beyond the pylorus extends the intestine with one or two loops to the anus. Connected with the intestine anteriorly is the large red mass of the liver, with its gall-bladder, which serves as a reservoir for bile, the fluid the liver secretes. Farther back is another red glandular mass, the spleen.
In front of the liver and separated from it by a membrane is the heart. This is of four parts. The posterior part is a thin-walled reservoir, the sinus venosus, into which blood enters through the jugular vein from the head and through the cardinal vein from the kidney. From the sinus venosus it passes forward into a large thin-walled chamber, the auricle.
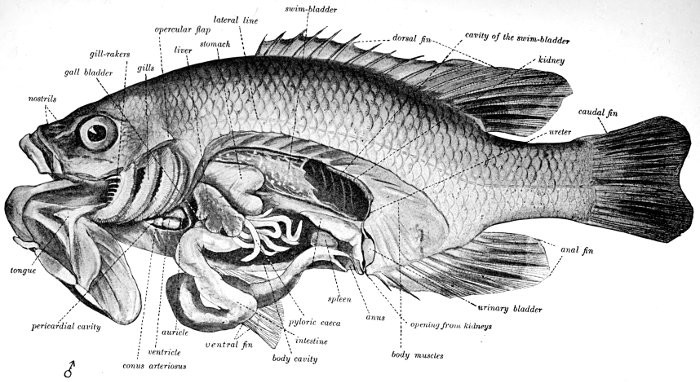
Fig. 16.—Dissection of the Blue-green Sunfish, Apomotis cyanellus Rafinesque. (After Kellogg.)—27.
Next it flows into the thick-walled ventricle, whence by the rhythmical contraction of its walls it is forced into an arterial bulb which lies at the base of the ventral aorta, which carries it on to the gills. After passing through the fine gill-filaments, it is returned to the dorsal aorta, a large blood-vessel which extends along the lower surface of the back-bone, giving out branches from time to time.
The kidneys in fishes constitute an irregular mass under the back-bone posteriorly. They discharge their secretions through the ureter to a small urinary bladder, and thence into the urogenital sinus, a small opening behind the anus. Into the same sinus are discharged the reproductive cells in both sexes.
In the female sunfish the ovaries consist of two granular masses of yellowish tissue lying just below and behind the swim-bladder. In the spring they fill much of the body cavity and the many little eggs can be plainly seen. When mature they are discharged through the oviduct to the urogenital sinus. In some fishes there is no special oviduct and the eggs pass into the abdominal cavity before exclusion.
In the male the reproductive organs have the same position as the ovaries in the female. They are, however, much smaller in size and paler in color, while the minute spermatozoa appear milky rather than granular on casual examination. A vas deferens leads from each of these organs into the urogenital sinus.
The lancelets, lampreys, and hagfishes possess no genital ducts. In the former the germ cells are shed into the atrial cavity, and from there find their way to the exterior either through the mouth or the atrial pore; in the latter they are shed directly into the body cavity, from which they escape through the abdominal pores. In the sharks and skates the Wolffian duct in the male, in addition to its function as an excretory duct, serves also as a passage for the sperm, the testes having a direct connection with the kidneys. In these forms there is a pair of Müllerian ducts which serve as oviducts in the females; they extend the length of the body cavity, and at their anterior end have an opening which receives the eggs which have escaped from the ovary into the body cavity. In some bony fishes as the eels and female salmon the germ cells are shed into the body cavity and escape through genital pores, which, however, may not be homologous with abdominal pores. In most other bony fishes the testes and ovaries are continued directly into ducts which open to the outside.
Organs of Nutrition.—The organs thus shown in dissection we may now examine in detail.
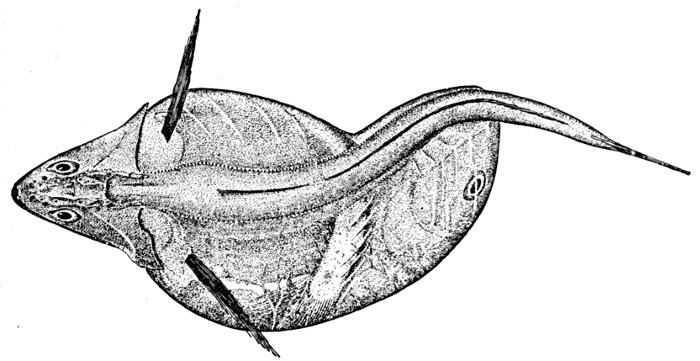
Fig. 17.—Black Swallower, Chiasmodon niger Johnson, containing a fish larger than itself. Le Have Bank.
The mouth of the fish is the organ or series of structures first concerned in nutrition. The teeth are outgrowths from the skin, primarily as modified papillæ, aiding the mouth in its various functions of seizing, holding, cutting, or crushing the various kinds of food material. Some fishes feed exclusively on plants, some on plants and animals alike, some exclusively on animals, some on the mud in which minute plants and animals occur. The majority of fishes feed on other fishes, and without much regard to species or condition. With the carnivorous fishes, to feed represents the chief activity of the organism. In proportion to the voracity of the fish is usually the size of the mouth, the sharpness of the teeth, and the length of the lower jaw.
The most usual type of teeth among fishes is that of villiform bands. Villiform teeth are short, slender, even, close-set, making a rough velvety surface. When the teeth are larger and more widely separated, they are called cardiform, like the teeth of a wool-card. Granular teeth are small, blunt, and sand-like. Canine teeth are those projecting above the level of the others, usually sharp, curved, and in some species barbed. Sometimes the canines are in front. In some families the last tooth in either jaw may be a "posterior canine," serving to hold small animals in place while the anterior teeth crush them. Canine teeth are often depressible, having a hinge at base.
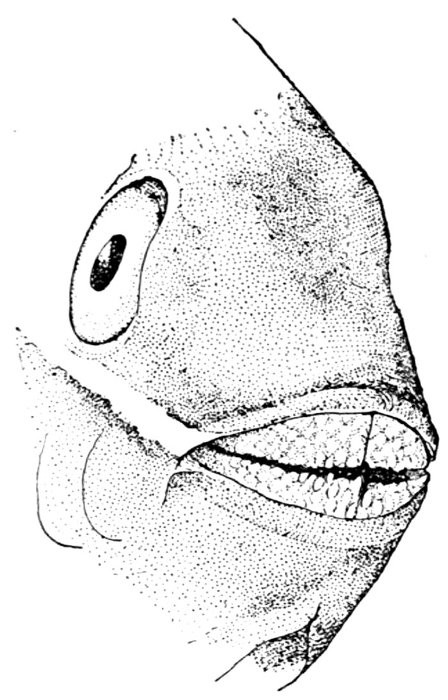
Fig. 18.—Jaws of a Parrot-fish, Sparisoma aurofrenatum (Val.). Cuba.
Teeth very slender and brush-like are called setiform. Teeth with blunt tips are molar. These are usually enlarged and fitted for crushing shells. Flat teeth set in mosaic, as in many rays and in the pharyngeals of parrot-fishes, are said to be paved or tessellated. Knife-like teeth, occasionally with serrated edges, are found in many sharks. Many fishes have incisor-like teeth, some flattened and truncate like human teeth, as in the sheepshead, sometimes with serrated edges. Often these teeth are movable, implanted only in the skin of the lips. In other cases they are set fast in the jaw. Most species with movable teeth or teeth with serrated edges are herbivorous, while strong incisors may indicate the choice of snails and crabs as food. Two or more of these different types may be found in the same fish. The knife-like teeth of the sharks are progressively shed, new ones being constantly formed on the inner margins of the jaw, so that the teeth are marching to be lost over the edge of the jaw as soon as each has fulfilled its function. In general the more distinctly a species is a fish-eater, the sharper are the teeth. Usually fishes show little discrimination in their choice of food; often they devour the young of their own species as readily as any other. The digestive process is rapid, and most fishes rapidly increase in size in the process of development. When food ceases to be abundant the fishes grow more slowly. For this reason the same species will grow to a larger size in large streams than in small ones, in lakes than in brooks. In most cases there is no absolute limit to growth, the species growing as long as it lives. But while some species endure many years, others are certainly very shortlived, and some may be even annual, dying after spawning, perhaps at the end of the first season.
Teeth are wholly absent in several groups of fishes. They are, however, usually present on the premaxillary, dentary, and pharyngeal bones. In the higher forms, the vomer, palatines, and gill-rakers are rarely without teeth, and in many cases the pterygoids, sphenoids, and the bones of the tongue are similarly armed.
No salivary glands or palatine velum are developed in fishes. The tongue is always bony or gristly and immovable. Sometimes taste-buds are developed on it, and sometimes these are found on the barbels outside the mouth.

Fig. 19.—Sheepshead (with incisor teeth), Archosargus probatocephalus (Walbaum). Beaufort, N. C.
The Alimentary Canal.—The mouth-cavity opens through the pharynx between the upper and lower pharyngeal bones into the œsophagus, whence the food passes into the stomach. The intestinal tract is in general divided into four portions—œsophagus, stomach, small and large intestines. But these divisions of the intestines are not always recognizable, and in the very lowest forms, as in the lancelet, the stomach is a simple straight tube without subdivision.
In the lampreys there is a distinction only of the œsophagus with many longitudinal folds and the intestine with but one. In the bony fishes the stomach is an enlarged area, either siphon-shaped, with an opening at either end, or else forming a blind sac with the openings for entrance (cardiac) and exit (pyloric) close together at the anterior end. In the various kinds of mullets (Mugil) and in the hickory shad (Dorosoma), fishes which feed on minute vegetation mixed with mud, the stomach becomes enlarged to a muscular gizzard, like that of a fowl. Attached near the pylorus and pouring their secretions into the duodenum or small intestine are the pyloric cæca. These are tubular sacs secreting a pale fluid and often almost as long as the stomach or as wide as the intestine. These may be very numerous as in the salmon, in which case they are likely to become coalescent at base, or they be few or altogether wanting.
Besides these appendages which are wanting in the higher vertebrates, a pancreas is also found in the sharks and many other fishes. This is a glandular mass behind the stomach, its duct leading into the duodenum and often coalescent with the bile duct from the liver. The liver in the lancelet is a long diverticulum of the intestine. In the true fishes it becomes a large gland of irregular form, and usually but not always provided with a gall-bladder as in the higher vertebrates. Its secretions usually pass through a ductus cholodechus to the duodenum.
The spleen, a dark-red lymphatic gland, is found attached to the stomach in all fish-like vertebrates except the lancelet.
The lining membrane of the abdominal cavity is known as the peritoneum, and the membrane sustaining the intestines from the dorsal side, as in the higher vertebrates, is called the mesentery. In many species the peritoneum is jet black, while in related forms it may be pale in color. It is more likely to be black in fishes from deep water and in fishes which feed on plants.
The Spiral Valve.—In the sharks or skates the rectum or large intestine is peculiarly modified, being provided with a spiral valve, with sometimes as many as forty gyrations. A spiral valve is also present in the more ancient types of the true fishes as dipnoans, crossopterygians, and ganoids. This valve greatly increases the surface of the intestine, doing away with the necessity for length. In the bowfin (Amia) and the garpike (Lepisosteus) the valve is reduced to a rudiment of three or four convolutions near the end of the intestine. In the sharks and skates the intestine opens into a cloaca, which contains also the urogenital openings. In all fishes the latter lie behind the orifice of the intestine. In the bony fishes and the ganoids there is no cloaca.

Fig. 20.—Stone-roller, Campostoma anomalum (Rafinesque). Family Cyprinidæ. Showing nuptial tubercles and intestines coiled about the air-bladder.
Length of the Intestine.—In all fishes, as in the higher vertebrates, the length of the alimentary canal is coordinated with the food of the fish. In those which feed upon plants the intestine is very long and much convoluted, while in those which feed on other fishes it is always relatively short. In the stone-roller, a fresh-water minnow (Campostoma) found in the Mississippi Valley, the excessively long intestines filled with vegetable matter are wound spool-fashion about the large air-bladder. In all other fishes the air-bladder lies on the dorsal side of the intestinal canal.
CHAPTER IV
THE SKELETON OF THE FISH

Specialization of the Skeleton.—In the lowest form of fish-like vertebrates (Branchiostoma), the skeleton consists merely of a cartilaginous rod or notochord extending through the body just below the spinal cord. In the lampreys, sharks, dipnoans, crossopterygians, and sturgeons the skeleton is still cartilaginous, but grows progressively more complex in their forms and relations. Among the typical fishes the skeleton becomes ossified and reaches a very high degree of complexity. Very great variations in the forms and relations of the different parts of the skeleton are found among the bony fishes, or teleostei. The high degree of specialization of these parts gives to the study of the bones great importance in the systematic arrangement of these fishes. In fact the true affinities of forms is better shown by the bones than by any other system of organs. In a general way the skeleton of the fish is homologous with that of man. The head in the one corresponds to the head in the other, the back-bone to the back-bone, and the paired fins, pectoral and ventral, to the arms and legs.
Homologies of Bones of Fishes.—But this homology does not extend to the details of structure. The bones of the arm of the specialized fish are not by any means identical with the humerus, coracoid, clavicle, radius, ulna, and carpus of the higher vertebrates. The vertebrate arm is not derived from the pectoral fin, but both from a cartilaginous shoulder-girdle with undifferentiated pectoral elements bearing fin-rays, in its details unlike an arm and unlike the pectoral fin of the specialized fish.
The assumption that each element in the shoulder-girdle and the pectoral fin of the fish must correspond in detail to the arm of man has led to great confusion in naming the different bones. Among the many bones of the fish's shoulder-girdle and pectoral fin, three or four different ones have successively borne the names of scapula, clavicle, coracoid, humerus, radius, and ulna. None of these terms, unless it be clavicle, ought by rights apply to the fish, for no bone of the fish is a true homologue of any of these as seen in man. The land vertebrates and the fishes have doubtless sprung from a common stock, but this stock, related to the crossopterygians of the present day, was unspecialized in the details of its skeleton, and from it the fishes and the higher vertebrates have developed the widely diverging lines.

Fig. 21.—Striped Bass, Roccus lineatus (Bloch). Potomac River.
Parts of the Skeleton.—The skeleton may be divided into the head, the vertebral column, and the limbs. The very lowest of the fish-like forms (Branchiostoma) has no differentiated head or skull, but in all the other forms the anterior part of the vertebral column is modified to form a cranium for the protection of the brain. In the lampreys there are no jaws or other appendages to the cranium.
In the sharks, dipnoans, crossopterygians, ganoids, and teleosts or bony fishes, jaws are developed as well as a variety of other bones around the mouth and throat. The jaw-bearing forms are sometimes known by the general name of gnathostomes. In the sharks and their relatives (rays, chimæras, etc.) all the skeleton is composed of cartilage. In the more specialized bony fishes, besides these bones we find also series of membrane bones, more or less external to the skull and composed of ossified dermal tissues. Membrane bones are not found in the sharks and lampreys, but are developed in an elaborate coat of mail in some extinct forms.
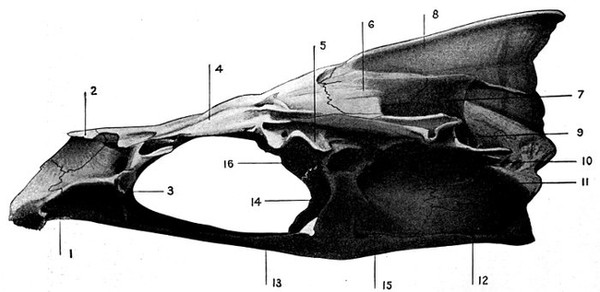
Fig. 22.—Roccus lineatus. Lateral view of cranium.
1. Vomer. 3. Prefrontal. 5. Sphenotic. 7. Epiotic. 9. Pterotic. 11. Exoccipital. 13. Parasphenoid. 15. Prootic. 2. Ethmoid. 4. Frontal. 6. Parietal. 8. Supraoccipital. 10. Opisthotic. 12. Basioccipital. 14. Basisphenoid.
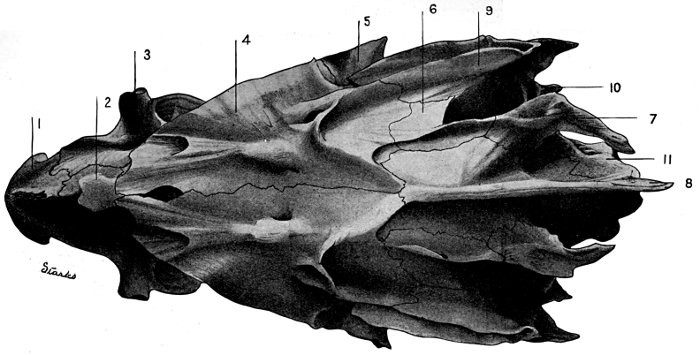
Fig. 23.—Roccus lineatus. Superior view of cranium.
1. Vomer. 3. Prefrontal. 5. Sphenotic. 7. Epiotic. 9. Pterotic. 11. Exoccipital. 2. Ethmoid. 4. Frontal. 6. Parietal. 8. Supraoccipital. 10. Opisthotic.
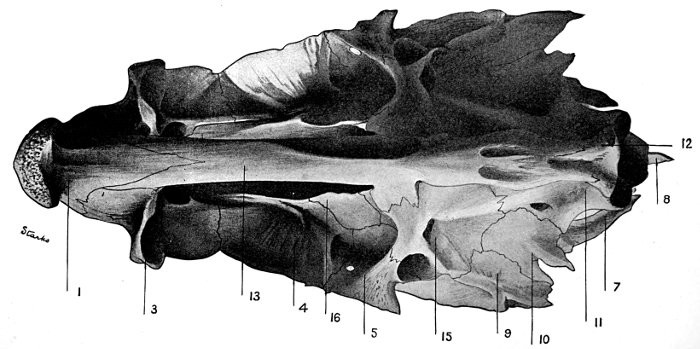
Fig. 24.—Roccus lineatus. Inferior view of cranium.
1. Vomer. 4. Frontal. 7. Epiotic. 9. Pterotic. 11. Exoccipital. 13. Parasphenoid. 16. Alisphenoid. 3. Prefrontal. 5. Sphenotic. 8. Supraoccipital. 10. Opisthotic. 12. Basioccipital. 15. Prootic.
Names of Bones of Fishes.—In the study of the names of the bones of fishes it will be more convenient to begin with a highly specialized form in which each of the various structures is present and in its normal position.
To this end we present a series of figures of a typical form, choosing, after Starks, the striped bass (Roccus lineatus) of the Atlantic coast of the United States. For this set of plates, drawn from nature by Mrs. Chloe Lesley Starks, we are indebted to the courtesy of Mr. Edwin Chapin Starks. The figures of the striped bass illustrate a noteworthy paper on "The Synonymy of the Fish Skeleton," published by the Washington Academy of Sciences in 1901.
Bones of the Cranium.—The vomer (1) is the anterior part of the roof of the mouth, armed with small teeth in the striped bass and in many other fishes, but often toothless. The ethmoid (2) lies behind the vomer on the upper surface of the skull, and the prefrontal (3) projects on either side and behind the ethmoid, the nostrils usually lying over or near it and near the nasal bone (51). Between the eyes above are the two frontal (4) bones joined by a suture. On the side behind the posterior angle of the frontal is the sphenotic (5) above the posterior part of the eye. Behind each frontal is the parietal (6). Behind the parietal and more or less turned inward over the ear-cavity is the epiotic (7). Between the parietals, and in most fishes rising into a thin crest, is the supraoccipital (8), which bounds the cranium above and behind, its posterior margin being usually a vertical knife-like edge. The pterotic (9) forms a sort of wing or free margin behind the epiotic and over the ear-cavity. The opisthotic (10) is a small, hard, irregular bone behind the pterotic. The exoccipital (11) forms a concave joint or condyle on each side of the basioccipital (12), by which the vertebral column is joined to the skull. The parasphenoid (13) forms a narrow ridge of the roof of the mouth, connecting the vomer with the basioccipital. In some fishes of primitive structure (Salmo, Beryx) there is another bone, called orbitosphenoid, on the middle line above and between the eyes. The basisphenoid (14) is a little bone above the myotome or tube in which runs the rectus muscle of the eye. It descends toward the parasphenoid and is attached to the prootic. The prootic (15) is an irregular bone below the ear region and lying in advance of the opisthotic. The alisphenoid (16) is a small bone in the roof of the mouth before the prootic. These sixteen bones (with a loose bone of specialized form, the otolith, within the ear-cavity) constitute the cranium. All are well developed in the striped bass and in most fishes. In some specialized forms they are much distorted, coossified, or otherwise altered, and their relations to each other may be more or less changed. In the lower forms they are not always fully differentiated, but in nearly all cases their homologies can be readily traced. In the sharks and lampreys the skull constitutes a continuous cartilaginous box without sutures. In the dipnoans and other forms having a bony casque the superficial bones outside the cranium may not correspond to the cartilaginous elements of the soft skull itself.
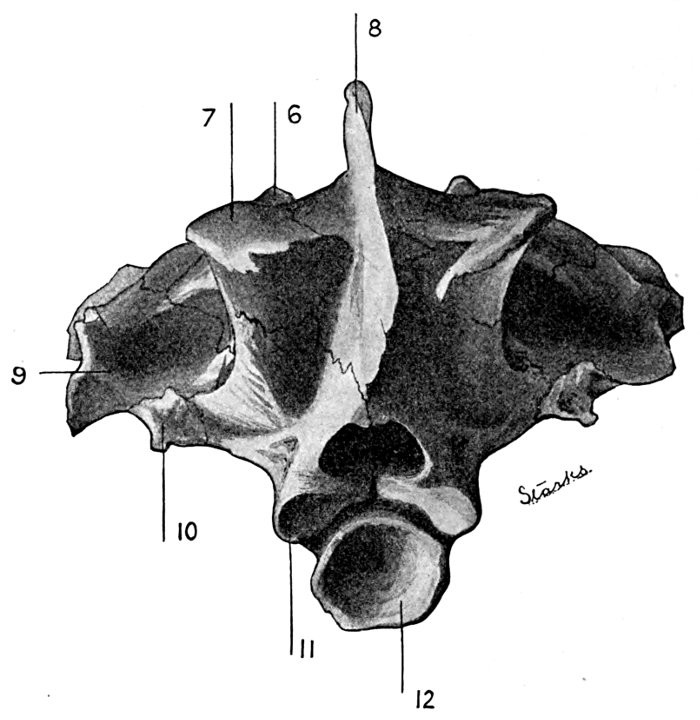
Fig. 25—Roccus lineatus. Posterior view of cranium.
- 6. Parietal.
- 7. Epiotic.
- 8. Supraoccipital.
- 9. Pterotic.
- 10. Opisthotic.
- 11. Exoccipital.
- 12. Basioccipital.
Bones of the Jaws.—The bones of the jaws are attached to the cranium by membranes only, not by sutures, except in a few peculiarly specialized forms.
The Upper Jaw.—The premaxillary (32) lies on either side and forms the front of the upper jaw. Its upper posterior tip or premaxillary spine projects backward almost at right angles with the rest of the bone into a groove on the ethmoid. There is often a fold in the skin by which this bone may be thrust out or protracted, as though drawn out of a sheath. When the spines of the premaxillary are very long the upper jaw may be thrust out for a considerable distance. The premaxillary is also often known as intermaxillary.
Lying behind the premaxillary, its anterior end attached within the angle of the premaxillary, is the maxillary (31), or supramaxillary, a flattened bone with expanded posterior tip. In the striped bass this bone is without teeth, but in many less specialized forms, as the salmon, it is provided with teeth and joined to the premaxillary in a different fashion. In any case its position readily distinguishes it. In some cases the maxillary is divided by one or more sutures, setting off from it one or more extra maxillary (supplemental maxillary) bones. This suture is absent in the striped bass, but distinct in the black bass, and more than one suture is found in the shad and herring. The roof of the mouth above is formed by a number of bones, which, as they often possess teeth, may be considered with the jaws. These are the palatine bones (21), one on either side flanking the vomer, the pterygoid (20), behind it and articulating with it, the mesopterygoid (22), on the roof of the mouth toward the median line, and the metapterygoid (23), lying behind this. Although often armed with teeth, these bones are to be considered of the general nature of the membrane bones. In some degraded types of fishes (eels, morays, congers) the premaxillary is indistinguishable, being united with the vomer and palatines.
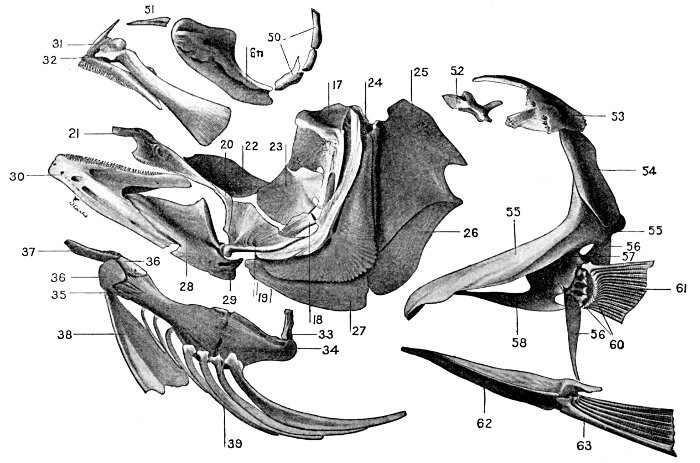
Fig. 26.—Roccus lineatus. Face-bones, shoulder and pelvic girdles, and hyoid arch.
- 17. Hyomandibular.
- 18. Symplectic.
- 19. Quadrate.
- 20. Pterygoid.
- 21. Palatine.
- 22. Mesopterygoid.
- 23. Metapterygoid.
- 24. Preopercle.
- 25. Opercle.
- 26. Subopercle.
- 27. Interopercle.
- 28. Articular.
- 29. Angular.
- 30. Dentary.
- 31. Maxillary.
- 32. Premaxillary.
- 33. Interhyal.
- 34. Epihyal.
- 35. Ceratohyal
- 36. Basihyal.
- 37. Glossohyal.
- 38. Urohyal.
- 39. Branchiostegal.
- 49. Preorbital.
- 50. Suborbital.
- 51. Nasal.
- 52. Supratemporal.
- 53. Post-temporal.
- 54. Supraclavicle.
- 55. Clavicle.
- 56. Postclavicle.
- 57. Hypercoracoid.
- 58. Hypocoracoid.
- 60. Actinosts.
- 61. Pectoral fin.
- 62. Pelvic girdle.
- 63. Ventral fin.
The upper jaw of the shark is formed from the anterior portion of the palatine bones, which are not separate from the quadrate, the whole forming the palato-quadrate apparatus. In the himæra and the dipnoans this apparatus is solidly united with the cranium. In these fishes the true upper jaw, formed of maxillary and premaxillary, is wanting.
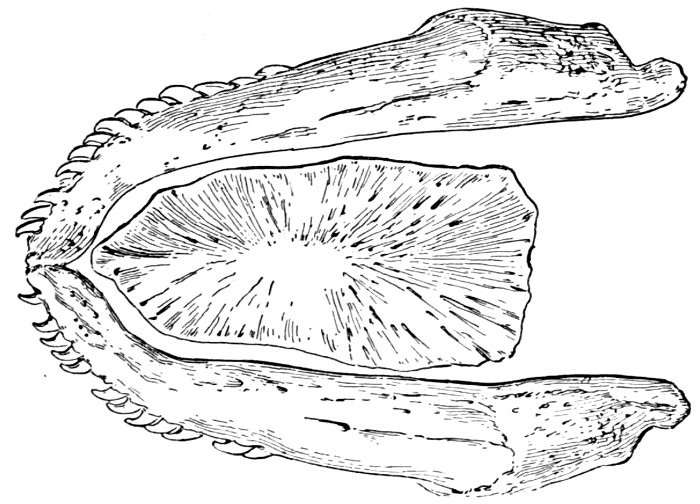
Fig. 27.—Lower jaw of Amia calva (Linnæus), showing the gular plate.
The Lower Jaw.—The lower jaw or mandible is also complex, consisting of two divisions or rami, right and left, joined in front by a suture. The anterior part of each ramus is formed by the dentary bone (30), which carries the teeth. Behind this is the articular bone (28), which is connected by a joint to the quadrate bone (19). At the lower angle of the articular bone is the small angular bone (29). In many cases another small bone, which is called splenial, may be found attached to the inner surface of the articular bone. This little bone has been called coronoid, but it is doubtless not homologous with the coronoid bone of reptiles. In a few fishes, Amia, Elopidæ, and certain fossil dipnoans, there is a bony gular plate, a membrane bone across the throat behind the chin on the lower jaw.
The Suspensorium of the Mandible.—The lower jaw is attached to the cranium by a chain of suspensory bones, which vary a good deal with different groups of fishes. The articular is jointed with the flat quadrate bone (19), which lies behind the pterygoid. A slender bone passes upward (18) under the preopercle and the metapterygoid, forming a connection above with a large flattish bone, the hyomandibular (17), which in turn joins the cranium. The slender bone which thus keys together the upper and lower elements, hyomandibular and quadrate, forming the suspensorium of the lower jaw, is known as symplectic (18). The hyomandibular is thought to be homologous with the stapes, or stirrup-bone, of the ear in higher animals. In this case the symplectic may be homologous with its small orbicular bone, and the malleus is a transformation of the articular. The incus, or anvil-bone, may be formed from part of Meckel's cartilage. All these homologies are however extremely hypothetical. The core of the lower jaw is formed of a cartilage called Meckel's cartilage, outside which the membrane bones, dentary, etc., are developed. This cartilage forms the lower jaw in sharks, true jaw-bones not being developed in these fishes. In lampreys and lancelets there is no lower jaw.
Membrane Bones of Face.—The membrane bones lie on the surface of the head, when they are usually covered by thin skin and have only a superficial connection with the cranium. Such bones, formed of ossified membrane, are not found in the earlier or less specialized fishes, the lancelets and lampreys, nor in the sharks, rays, and chimæras. They are chiefly characteristic of the bony fishes, although in some of these they have undergone degradation.
The preorbital (49) lies before and below the eye, its edge more or less parallel with that of the maxillary. It may be broad or narrow. When broad it usually forms a sheath into which the maxillary slips. The nasal (51) lies before the preorbital, a small bone usually lying along the spine of the premaxillary. Behind and below the eye is a series of about three flat bones, the suborbitals (50), small in the striped bass, but sometimes considerably modified. In the great group of loricate fishes (sculpins, etc.), the third suborbital sends a bony process called the suborbital stay backward across the cheek toward the preopercle. The suborbital stay is present in the rosefish. In some cases, as in the gurnard, this stay covers the whole cheek with a bony coat of mail. In some fishes, but not in the striped bass, a small supraorbital bone exists over the eye, forming a sort of cap on an angle of the frontal bone.
The largest uppermost flat bone of the gill-covers is known as the opercle (25). Below it, joined by a suture, is the subopercle (26). Before it is the prominent ridge of the preopercle (24), which curves forward below and forms a more or less distinct angle, often armed with serrations or spines. In some cases this armature is very highly developed. The interopercle (27) lies below the preopercle and parallel with the lower limb.
Branchial Bones.—The bones of the branchial apparatus or gills are very numerous and complex, as well as subject to important variations. In many fishes some of these bones are coossified, and in other cases some are wanting. The tongue may be considered as belonging to this series, as the bones of the gills are attached to its axis below.
In the striped bass, as in most fishes, the tongue, gristly and immovable, is formed anteriorly by a bone called the glossohyal (37). Behind this are the basihyals (36), and still farther back, on the side, is the ceratohyal (35). To the basihyals is attached a bone extending downward and free behind the urohyal (38). Behind the ceratohyal and continuous with it is the epihyal (34), to which behind is attached the narrow interhyal (33). On the under surface of the ceratohyal and the epihyal are attached the branchiostegals (39). These are slender rays supporting a membrane beneath the gills, seven in number on each side in the striped bass, but much more numerous in some groups of fishes. The gill membranes connecting the branchiostegals are in the striped bass entirely separate from each other. In other fishes they may be broadly joined across the fleshy interspace between the gill-openings, known as the isthmus, or again they may be grown fast to the isthmus itself, so that the gill-openings of the two sides are widely separated.
The Gill-arches.—The gills are attached to four bony arches with a fifth of the same nature, but totally modified by the presence of teeth, and very rarely having on it any of the gill-fringes. The fifth arch thus modified to serve in mastication instead of respiration is known collectively as the lower pharyngeals (46). Opposite these are the upper pharyngeals (45).
The gill-arches are suspended to the cranium from above by the suspensory pharyngeal (44). Each arch contains three parts—the epibranchial (43), above, the ceratobranchial (42), forming the middle part, and the hypobranchial (41), the lower part articulating with the series of basibranchials (40) which lie behind the epihyal of the tongue. On the three bones forming the first gill-arch are attached numerous appendages called gill-rakers (47). These gill-rakers vary very greatly in number and form. In the striped bass they are few and spear-shaped. In the shad they are very many and almost as fine as hairs. In some fishes they form an effective strainer in separating the food, or perhaps in keeping extraneous matter from the gills. In some fishes they are short and lumpy, in others wanting altogether.
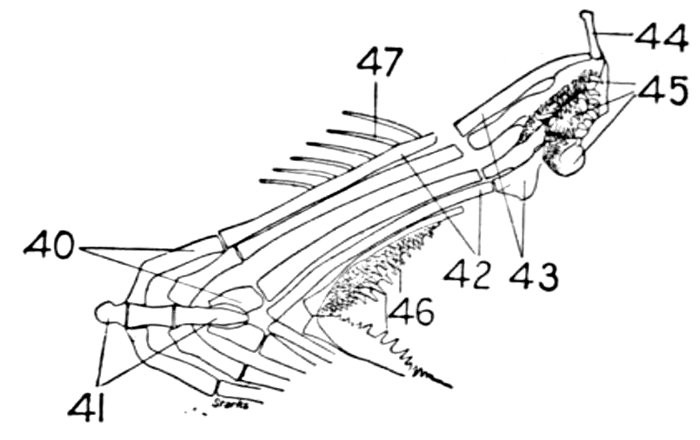
Fig. 28.—Roccus lineatus. Branchial arches. (After Starks.)
- 40. Basibranchial.
- 41. Hypobranchial.
- 42. Ceratobranchial.
- 43. Epibranchial.
- 44. Suspensory pharyngeal.
- 45. Upper pharyngeals.
- 46. Lower pharyngeals.
- 47. Gill-rakers.
The Pharyngeals.—The hindmost gill-arch, as above stated, is modified to form a sort of jaw. The tooth-bearing bones above, 2 to 4 pairs, are known as upper pharyngeals (45), those below, single pair, as lower pharyngeals (46). Of these the lower pharyngeals are most highly specialized and the most useful in classification. These are usually formed much as in the striped bass. Occasionally they are much enlarged, with large teeth for grinding. In many families the lower pharyngeals are grown together in one large bone. In the suckers (Catostomidæ) the lower pharyngeal preserves its resemblance to a gill-arch. In the carp family (Cyprinidæ) retaining this resemblance, it possesses highly specialized teeth.
Vertebral Column.—The vertebral column is composed of a series of vertebræ, 24 in number in the striped bass and in many of the higher fishes, but varying in different groups from 16 to 18 to upwards of 400, the higher numbers being evidence of unspecialized or more usually degenerate structure.
Each vertebra consists of a double concave body or centrum (66). Above it are two small projections often turned backward, zygapophyses (71), and two larger ones, neurapophyses (67), which join above to form the neural spine (68) and thus form the neural canal, through which passes the spinal cord from end to end of the body.
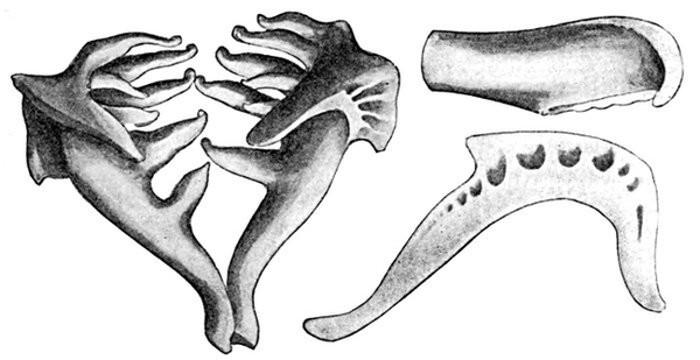
Fig. 29.—Pharyngeal bone and teeth of European Chub, Leuciscus cephalus (Linnæus). (After Seelye.)
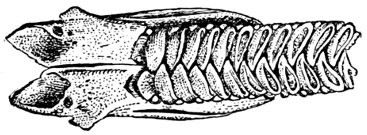
Fig. 30.—Upper pharyngeals of a Parrot-fish, Scarus strongylocephalus.
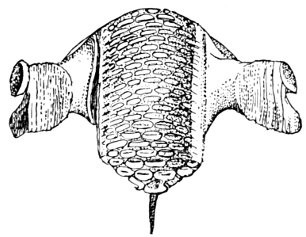
Fig. 31.—Lower pharyngeals of a Parrot-fish, Scarus strongylocephalus (Bleeker).
Below in the vertebræ of the posterior half of the body the hæmapophyses (69) unite to form the hæmal spine (70), and through the hæmal canal thus formed passes a great artery. The vertebræ having hæmal as well as neural spines are known as caudal vertebræ, and occupy the posterior part of the body, usually that behind the attachment of the anal fin (78).
The anterior vertebræ known as abdominal vertebræ, bounding the body-cavity, possess neural spines similar to those of the caudal vertebræ. In place, however, of the hæmapophyses are projections known as parapophyses (72), which do not meet below, but extend outward, forming the upper part of the wall of the abdominal cavity.

Fig. 32.—Pharyngeals of Italian Parrot-fish, Sparisoma cretense (L.). a, upper; b, lower.
To the parapophyses, or near them, the ribs (73) are rather loosely attached and each rib may have one or more accessory branches (74) called epipleurals.
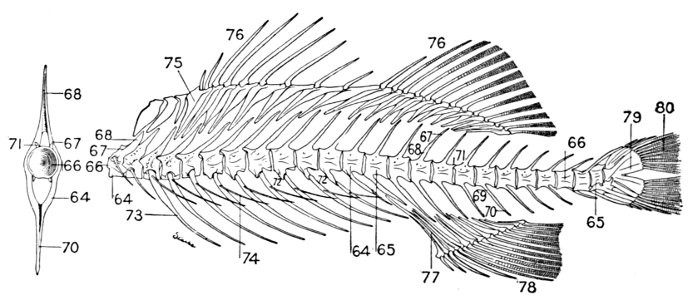
Fig. 33.—Roccus lineatus. Vertebral column and appendages, with a typical vertebra. (After Starks.)
- 64. Abdominal vertebræ.
- 65. Caudal vertebræ.
- 66. Centrum.
- 67. Neurapophysis.
- 68. Neural spine.
- 69. Hæmapophysis.
- 70. Hæmal spine.
- 71. Zygapophysis.
- 72. Parapophysis.
- 73. Ribs.
- 74. Epipleurals.
- 75. Interneural.
- 76. Dorsal fin.
- 77. Interhæmal.
- 78. Anal fin.
- 79. Hypural.
- 80. Caudal fin.
In the striped bass the dorsal vertebræ are essentially similar in form, but in some fishes, as the carp and the catfish, 4 or 5 anterior vertebræ are greatly modified, coossified, and so arranged as to connect the air-bladder with the organ of hearing. Fishes with vertebræ thus altered are called plectospondylous.
In the garpike the vertebræ are convex anteriorly, concave behind, being joined by ball-and-socket joints (opisthocœlian). In most other fishes they are double concave (amplicœlian). In sharks the vertebræ are imperfectly ossified, a number of terms, asterospondylous, cyclospondylous, tectospondylous, being applied to the different stages of ossification, these terms referring to the different modes of arrangement of the calcareous material within the vertebra.
The Interneurals and Interhæmals.—The vertical fins are connected with the skeletons by bones placed loosely in the flesh and not joined by ligament or suture. Below the dorsal fin (76) lies a series of these bones, dagger-shaped, with the point downward. These are called interneurals (75) and to these the spines and soft rays of the fin are articulated.
In like fashion the spines and rays of the anal fin (18) are jointed at base to bones called interhæmals (77). In certain cases the second interhæmal is much enlarged, made hollow and quill-shaped, and in its concave upper end the tip of the air-bladder is received. This structure is seen in the plume-fishes (Calamus). These two groups of bones, interneural and interhæmal, are sometimes collectively called inter-spinals. The flattened basal bone of the caudal fin (80) is known as hypural (79).

Fig. 34.—Basal bone of dorsal fin, Holoptychius leptopterus (Agassiz). (After Woodward.)
The tail of the striped bass, ending in a broad plate which supports the caudal, is said to be homocercal. In more primitive forms the tail is turned upward more or less, the fin being largely thrown to its lower side. Such a tail as in the sturgeon is said to be heterocercal. In the isocercal tail of the codfish and its relatives the vertebræ are progressively smaller behind and the hypural plate is obsolete or nearly so, the vertebræ remaining in the line of the axis of the body and dividing the caudal fin equally. The simplest form of tail, called diphycercal, is extended horizontally, tapering backward, the fin equally divided above and below, without hypural plate. In any form of the tail, it may through degeneration be attenuate or whip-like, a form called leptocercal.
The Pectoral Limb.—The four limbs of the fish are represented by the paired fins. The anterior limb is represented by the pectoral fin and its basal elements with the shoulder-girdle, which in the bony fishes reaches a higher degree of complexity than in any other vertebrates. It is in connection with the shoulder-girdle that the greatest confusion in names has occurred. This is due to an attempt to homologize its parts with the shoulder-girdle (scapula, coracoid, and clavicle) of higher vertebrates. But it is not evident that a bony fish possesses a real scapula, coracoid, or even clavicle. The parts of its shoulder-girdle are derived by one line of descent from the undifferentiated elements of the cartilaginous shoulder-girdle of ancestral crossopterygian or dipnoan forms. From a similar ancestry by another line of differentiation has come the amphibian and reptilian shoulder-girdle and its derivative, the girdle of birds and mammals.
The Shoulder-girdle.—In the higher fishes the uppermost bone of the shoulder-girdle is called the post-temporal (suprascapula) (53). In the striped bass and in most fishes this bone is jointed to the temporal region of the cranium. Sometimes, as in the trigger-fishes, it is grown fast to the skull, but it usually rests lightly with the three points of its upper end. In sharks and skates the shoulder-girdle, which is formed of a continuous cartilage, does not touch the skull. In the eels and their allies, it has, by degradation, lost its connection and the post-temporal rests in the flesh behind the cranium.
The post-temporal sometimes projects behind through the skin and may bear spines or serrations. In front of the post-temporal and a little to the outside of it is the small supratemporal (52) also usually connecting the shoulder-girdle with the skull. Below the post-temporal, extending downward and backward, is the flattish supraclavicle (posterotemporal) (54). To this is joined the long clavicle (proscapula) (55), which runs forward and downward in the bony fishes, meeting its fellow on the opposite side in a manner suggesting the wishbone of a fowl. Behind the base of the clavicle, the sword-shaped post-clavicle (56) extends downward through the muscles behind the base of the pectoral fin. In some fishes, as the stickleback and the trumpet-fish, a pair of flattish or elongate bones called interclavicles (infraclavicles) lie between and behind the lower part of the clavicle. These are not found in most fishes and are wanting in the striped bass. They are probably in all cases merely extensions of the hypocoracoid.
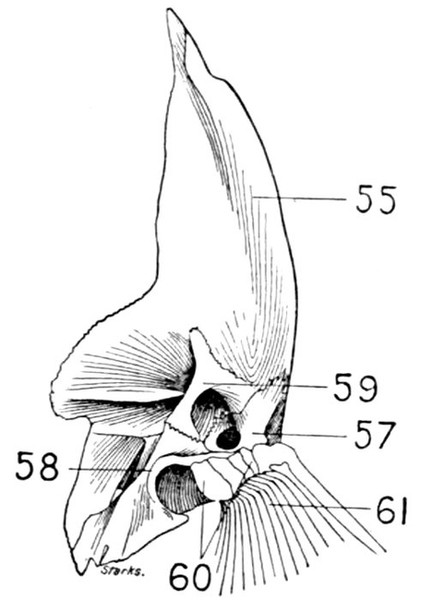
Fig. 35.—Inner view of shoulder-girdle of the Buffalo-fish, Ictiobus bubalus Rafinesque, showing the mesocoracoid (59). (After Starks.)
Two flat bones side by side lie at the base of the pectoral fin, their anterior edges against the upper part of the clavicle. These are the hypercoracoid (57), above, and hypocoracoid (58), below. These have been variously called scapula, coracoid, humerus, radius, and ulna, but being found in the higher fishes only and not in the higher vertebrates, they should receive names not used for other structures. The hypercoracoid is usually pierced by a round foramen or fenestra, but in some fishes (cods, weavers) the fenestra is between the two bones. Attached to the hypercoracoid in the striped bass are four little bones shaped like an hour-glass. These are the actinosts (60) (carpals or pterygials), which support the rays of the pectoral fin (61). In most bony fishes these are placed much as in the striped bass, but in certain specialized or aberrant forms their form and position are greatly altered.
In the anglers (Pediculati) the "carpals" are much elongated, forming a kind of arm, by which the fish can execute a motion not unlike walking.
In the Alaska blackfish (Dallia pectoralis) the two coracoids are represented by a thin, cartilaginous plate, imperfectly divided, and there are no actinosts. In almost all bony fishes, however, these bones are well differentiated and distinct. In most of the soft-rayed fishes an additional V-shaped bone or arch exists on the inner surface of the shoulder-girdle near the insertion of the hypercoracoid. This is known as the mesocoracoid (59). It is not found in the striped bass, but is found in the carp, catfish, salmon, and all their allies.

Fig. 36.—Sargassum-fish, Pterophryne tumida (Osbeck). One of the Anglers. Family Antennariidæ.
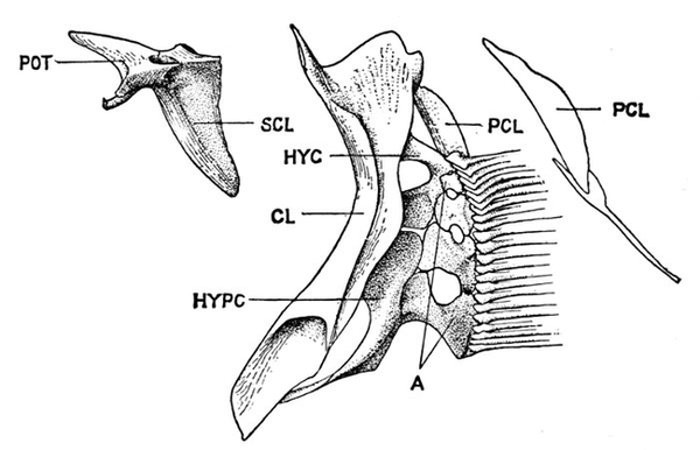
Fig. 37.—Shoulder-girdle of Sebastolobus alascanus Gilbert. (After Starks.)
- POT. Post-temporal.
- CL. Clavicle.
- PCL. Postclavicle.
- HYC. Hypercoracoid.
- HYPC. Hypocoracoid.
The Posterior Limbs.—The posterior limb or ventral fin (63) is articulated to a single bone on either side, the pelvic girdle (62).
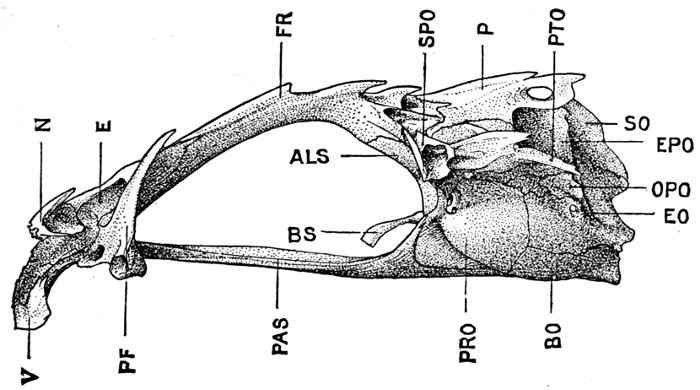
Fig. 38.—Cranium of Sebastolobus alascanus Gilbert. (After Starks.)
- V. Vomer.
- N. Nasal.
- E. Ethmoid.
- PF. Prefrontal.
- FR. Frontal.
- PAS. Parasphenoid.
- ALS. Alisphenoid.
- P. Parietal.
- BA. Basisphenoid.
- PRO. Prootic.
- BO. Basioccipital.
- SO. Supraoccipital.
- EO. Exoccipital.
- EPO. Epiotic.
- SPO. Sphenotic.
- PTO. Pterotic.
In the shark the pelvic girdle is rather largely developed, but in the more specialized fishes it loses its importance. In the less specialized of the bony fishes the pelvis is attached at a distance from the head among the muscles of the side, and free from the shoulder-girdle and other parts of the skeleton. The ventral fins are then said to be abdominal. When very close to the clavicle, but not connected with it, as in the mullet, the fin is still said to be abdominal or subabdominal. In the striped bass the pelvis is joined by ligament between the clavicles, near their tip. The ventral fins thus connected, as seen in most spiny-rayed fishes, are said to be thoracic. In certain forms the pelvis is thrown still farther forward and attached at the throat or even to the chin. When the ventral fins are thus inserted before the shoulder-girdle, they are said to be jugular. Most of the fishes with spines in the fins have thoracic ventrals. In the fishes with jugular ventrals these fins have begun a process of degeneration by which the spines or soft rays or both are lost or atrophied.
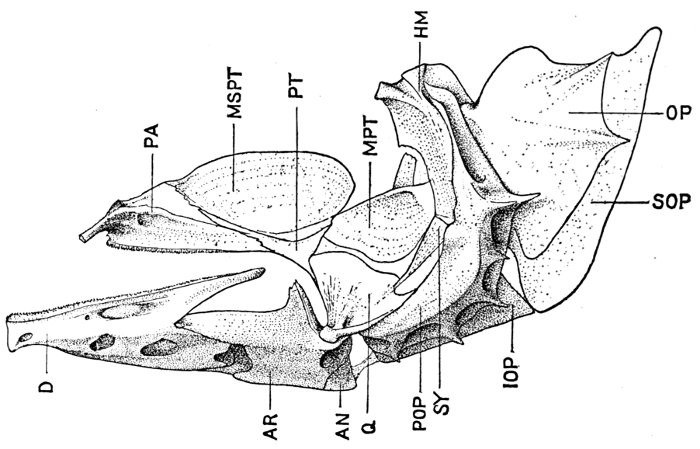
Fig. 39.—Lower jaw and palate of Sebastolobus alascanus. (After Starks.)
- PA. Palatine.
- MSPT. Mesopterygoid.
- PT. Pterygoid.
- MPT. Metapterygoid.
- D. Dentary.
- AR. Articular.
- AN. Angular.
- Q. Quadrate.
- SY. Symplectic.
- HM. Hyomandibular.
- POP. Preopercle.
- IOP. Interopercle.
- SOP. Subopercle.
- OP. Opercle.
Degeneration.—By degeneration or degradation in biology is meant merely a reduction to a lower degree of complexity or specialization in structure. If in the process of development of the individual some particular organ loses its complexity it is said to be degenerate. If in the geological history of a type the same change takes place the same term is used. Degeneration in this sense is, like specialization, a phase of adaptation. It does not imply disease, feebleness, or mutilation, or any tendency toward extinction. It is also necessary to distinguish clearly phases of primitive simplicity from the apparent simplicity resulting from degeneration.
The Skeleton in Primitive Fishes.—To learn the names of bones we can deal most satisfactorily with the higher fishes, those in which the bony framework has attained completion. But to understand the origin and relation of parts we must begin with the lowest types, tracing the different stages in the development of each part of the system.
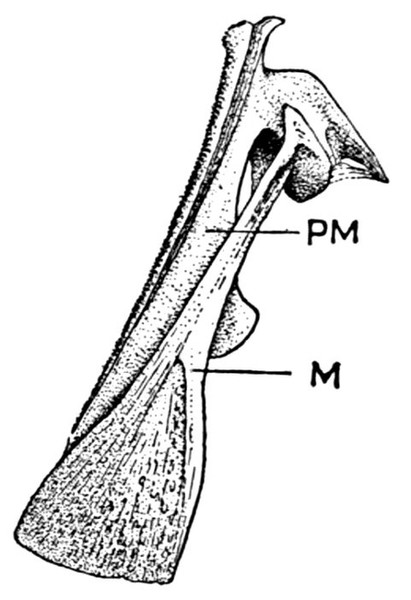
Fig. 40.—Maxillary and premaxillary of Sebastolobus alascanus. M, maxillary; PM, premaxillary.
In the lancelets (Leptocardii), the vertebral column consists simply of a gelatinous notochord extending from one end of the fish to the other, and pointed at both ends, no skull being developed. The notochord never shows traces of segmentation, although cartilaginous rods above it are thought to forecast apophyses. In these forms there is no trace of jaws, limbs, or ribs.
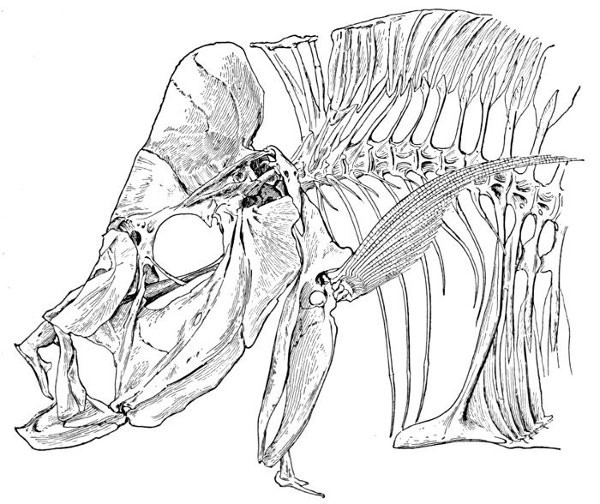
Fig. 41.—Part of skeleton of Selene vomer (Linnæus).
In the embryo of the bony fish a similar notochord precedes the segmentation and ossification of the vertebral column. In most of the extinct types of fishes a notochord more or less modified persisted through life, the vertebræ being strung upon it spool fashion in various stages of development. In the Cyclostomi (lampreys and hagfishes) the limbs and lower jaw are still wanting, but a distinct skull is developed. The notochord is still present, but its anterior pointed end is wedged into the base of a cranial capsule, partly membranous, partly cartilaginous. There is no trace of segmentation in the notochord itself in these or any other fishes, but neutral arches are foreshadowed in a series of cartilages on each side of the spinal chord. The top of the head is protected by broad plates. There are ring-like cartilages supporting the mouth and other cartilages in connection with the tongue and gill structures.
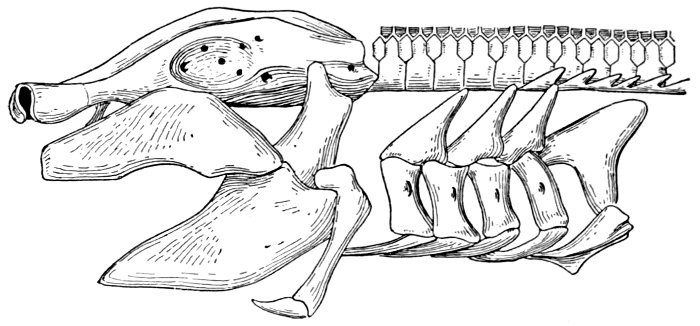
Fig. 42.—Hyostylic skull of Chiloscyllium indicum, a Scyliorhinoid Shark. (After Parker and Haswell.)
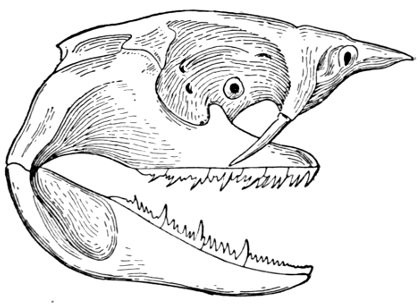
Fig. 43.—Skull of Heptranchias indicus (Gmelin), a notidanoid shark. (After Parker and Haswell.)
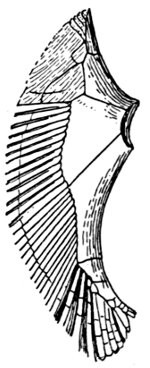
Fig. 44.—Basal bones of pectoral fin of Monkfish, Squatina. (After Zittel.)
The Skeleton of Sharks.—In the Elasmobranchs (sharks, rays, chimæras) the tissues surrounding the notochord are segmented and in most forms distinct vertebræ are developed. Each of these has a conical cavity before and behind, with a central canal through which the notochord is continued. The form and degree of ossification of these vertebræ differ materially in the different groups. The skull in all these fishes is cartilaginous, forming a continuous undivided box containing the brain and lodging the organs of sense. To the skull in the shark is attached a suspensorium of one or two pieces supporting the mandible and the hyoid structures. In the chimæra the mandible is articulated directly with the skull, the hyomandibular and quadrate elements being fused with the cranium. The skull in such case is said to be autostylic, that is, with self-attached mandible. In the shark it is said to be hyostylic, the hyomandibular intervening. The upper jaw in the shark consists not of maxillary and premaxillary but of palatine elements, and the two halves of the lower jaw are representatives of Meckel's cartilage, which is the cartilaginous centre of the dentary bone in the bony fishes. These jaw-bones in the higher fishes are in the nature of membrane bones, and in the sharks and their relatives all such bones are undeveloped. The hyoid structures are in the shark relatively simple, as are also the gill-arches, which vary in number. The vertical fins are supported by interneural and interhæmal cartilages, to which the soft fin-rays are attached without articulation.
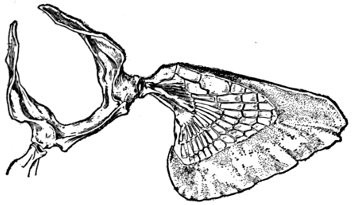
Fig. 45.—Pectoral fin of Heterodontus philippi. (From nature.)
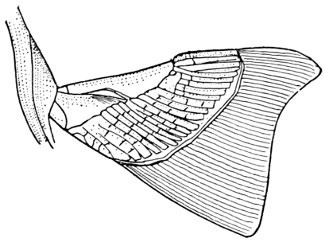
Fig. 46.—Pectoral fin of Heptranchias indicus (Gmelin). (After Dean.)
The shoulder-girdle is made of a single cartilage, touching the back-bone at a distance behind the head. To this cartilage three smaller ones are attached, forming the base of the pectoral fin. These are called mesopterygium, propterygium, and metapterygium, the first named being in the middle and more distinctly basal. These three segments are subject to much variation. Sometimes one of them is wanting; sometimes two are grown together. Behind these the fin-rays are attached. In most of the skates the shoulder-girdle is more closely connected with the anterior vertebræ, which are more or less fused together.
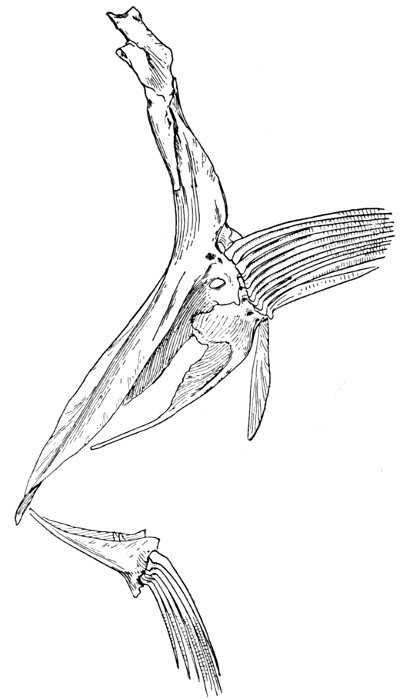
Fig. 47.—Shoulder-girdle of a Flounder, Paralichthys californicus (Ayres).
The pelvis, remote from the head, is formed, in the shark, of a single or paired cartilage with smaller elements at the base of the fin-rays. In the males a cartilaginous generative organ, known as the clasper, is attached to the pelvis and the ventral fins. In the Elasmobranchs the tail vertebræ are progressively smaller backward. If a caudal fin is present, the last vertebræ are directed upward (heterocercal) and the greater part of the fin is below the axis. In other forms (sting-rays) the tail degenerates into a whip-like organ (leptocercal), often without fins. In certain primitive sharks (Ichthyotomi), as well as in the Dipnoi and Crossopterygii, the tail is diphycercal, the vertebræ growing progressively smaller backward and not bent upward toward the tip.
In the chimæras (Holocephali) the notochord persists and is surrounded by a series of calcified rings. The palate with the suspensorium is coalesced with the skull, and the teeth are grown together into bony plates.
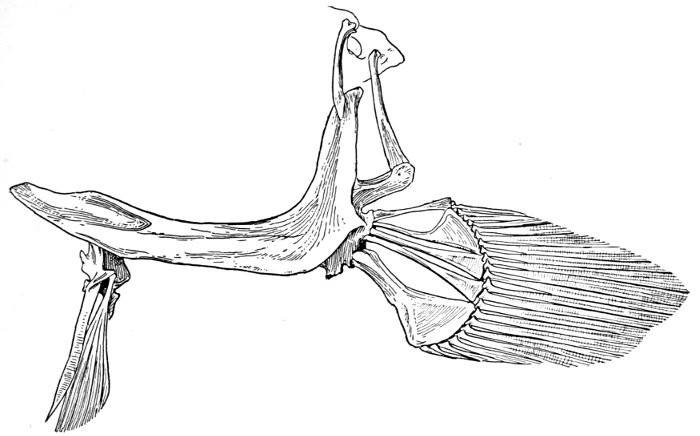
Fig. 48.—Shoulder-girdle of a Toadfish, Batrachoides pacifici (Günther).
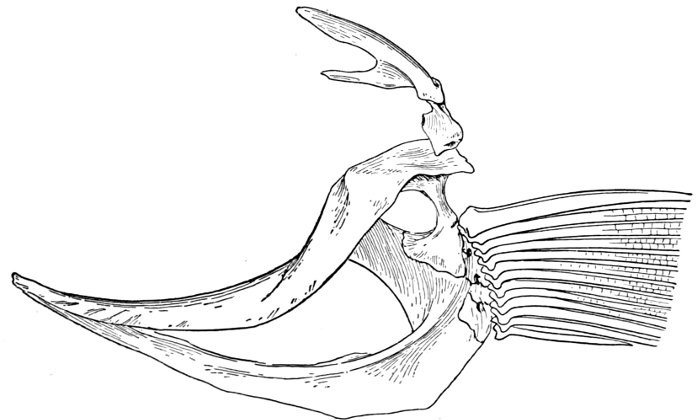
Fig. 49.—Shoulder-girdle of a Garfish, Tylosurus fodiator (Jordan and Gilbert).
The Archipterygium.—The Dipnoans, Crossopterygians, and Ganoids represent various phases of transition from the ancient cartilaginous types to the modern bony fishes.
In the Ichthyotomous sharks, Dipnoans, and Crossopterygians the segments of the pectoral limb are arranged axially, or one beyond another. This type of fin has been called archipterygium by Gegenbaur, on the theory that it represents the condition shown on the first appearance of the pectoral fin. This theory is now seriously questioned, but it will be convenient to retain the name for the pectoral fin with segmented axis fringed on one or both sides by soft rays.
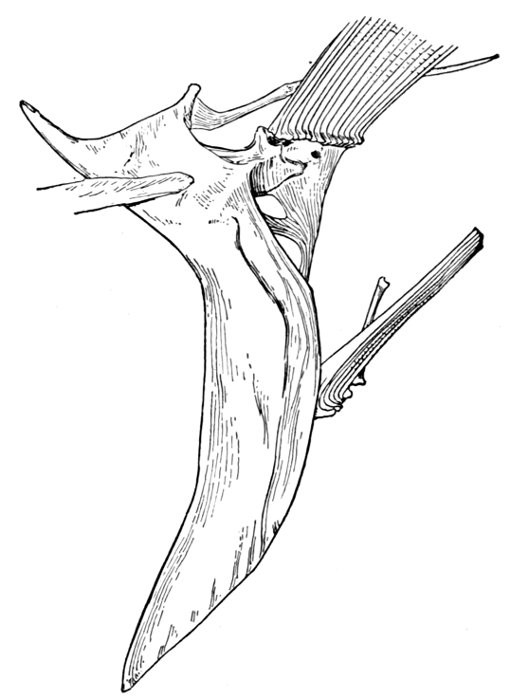
Fig. 50.—Shoulder-girdle of a Hake, Merluccius productus (Ayres).
The archipterygium of the Dipnoan genus Neoceratodus is thus described by Dr. Günther ("Guide to the Study of Fishes," p. 73): "The pectoral limb is covered with small scales along the middle from the root to the extremity, and is surrounded by a rayed fringe similar to the rays of the vertical fins. A muscle split into numerous fascicles extends all the length of the fin, which is flexible in every part and in every direction. The cartilaginous framework supporting it is joined to the scapular arch by a broad basal cartilage, generally single, sometimes showing traces of a triple division. Along the middle of the fin runs a jointed axis gradually becoming smaller and thinner towards the extremity. Each joint bears on each side a three-, two-, or one-jointed branch."
In the genus Lepidosiren, also a Dipnoan, the pectoral limb has the same axial structure, but is without fin-rays, although in the breeding season the posterior limb or ventral fin in the male is covered with a brush of fine filaments. This structure, according to Prof. J. G. Kerr,[3] is probably without definite function, but belongs to the "category of modifications so often associated with the breeding season (cf. the newts' crest) commonly called ornamental, but which are perhaps more plausibly looked upon as expressions of the intense vital activity of the organisms correlated with its period of reproductive activity." Professor Kerr, however, thinks it not unlikely that this brush of filaments with its rich blood-supply may serve in the function of respiration, a suggestion first made by Professor Lankester.
FOOTNOTES:
[3] Philos. Trans., Lond., 1900.
CHAPTER V
MORPHOLOGY OF THE FINS

Origin of the Fins of Fishes.—One of the most interesting problems in vertebrate morphology, and one of the most important from its wide-reaching relations, is that of the derivation of the fins of fishes. This resolves itself at once into two problems, the origin of the median fins, which appear in the lancelets, at the very bottom of the fish-like series, and the origin of the paired fins or limbs, which are much more complex, and which first appear with the primitive sharks.
In this study the problem is to ascertain not what theoretically should happen, but what, as a matter of fact, has happened in the early history of the fish-like groups. That these structures, with the others in the fish body, have sprung from simple origins, growing more complex with the demands of varied conditions, and then at times again simple, through degeneration, there can be no doubt. It is also certain that each structure must have had some element of usefulness in all its stages. In such studies we have, as Hæckel has expressed it, "three ancestral documents, paleontology, morphology, and ontogeny"—the actual history as shown by fossil remains, the sidelight derived from comparison of structures, and the evidence of the hereditary influences shown in the development of the individual. As to the first of these ancestral documents, the evidence of paleontology is conclusive where it is complete. But in very few cases are we sure of any series of details. The records of geology are like a book with half its leaves torn out, the other half confused, displaced, and blotted. Still each record actually existing represents genuine history, and in paleontology we must in time find our final court of appeal in all matters of biological origins.
The evidence of comparative anatomy is most completely secured, but it is often indecisive as to relative age and primitiveness of origin among structures. As to ontogeny, it is, of course, true that through heredity "the life-history of the individual is an epitome of the life-history of the race." "Ontogeny repeats phylogeny," and phylogeny, or line of descent of organisms and structures, is what we are seeking. But here the repetition is never perfect, never nearly so perfect in fact as Hæckel and his followers expected to find it. The demands of natural selection may lead to the lengthening, shortening, or distortion of phases of growth, just as they may modify adult conditions. The interpolation of non-ancestral stages is recognized in several groups. The conditions of the individual development may, therefore, furnish evidence in favor of certain theories of origins, but they cannot alone furnish the absolute proof.
In the process of development the median or vertical fins are doubtless older than the paired fins or limbs, whatever be the origin of the latter. They arise in a dermal keel which is developed in a web fitting and accentuating the undulatory motion of the body. In the embryo of the fish the continuous vertical fin from the head along the back and around the tail precedes any trace of the paired fins.
In this elementary fin-fold slender supports, the rudiments of fin-rays, tend to appear at intervals. These are called by Ryder ray-hairs or actinotrichia. They are the prototype of fin-rays in the embryo fish, and doubtless similarly preceded the latter in geological time. In the development of fishes the caudal fin becomes more and more the seat of propulsion. The fin-rays are strengthened, their basal supports are more and more specialized, and the fin-fold ultimately divides into distinct fins, the longest rays developed where most needed.
That the vertical fins, dorsal, anal, and caudal, have their origin in a median fold of the skin admits of no question. In the lowest forms which bear fins these structures are dermal folds, being supported by very feeble rays. Doubtless at first the vertical fins formed a continuous fold, extending around the tail, this fold ultimately broken, by atrophy of parts not needed, into distinct dorsal, anal, and caudal fins. In the lower fishes, as in the earlier sharks, there is an approach to this condition of primitive continuity, and in the embryos of almost all fishes the same condition occurs. Dr. John A. Ryder points out the fact that there are certain unexplained exceptions to this rule. The sea-horse, pipefish, and other highly modified forms do not show this unbroken fold, and it is wanting in the embryo of the top-minnow, Gambusia affinis. Nevertheless the existence of a continuous vertical fold in the embryo is the rule, almost universal. The codfish with three dorsals, the Spanish mackerel with dorsal and anal finlets, the herring with one dorsal, the stickleback with a highly modified one, all show this character, and we may well regard it as a certain trait of the primitive fish. This fold springs from the ectoblast or external series of cells in the embryo. The fin-rays and bony supports of the fins spring from the mesoblast or middle series of cells, being thrust upward from the skeleton as supports for the fin-fold.
Origin of the Paired Fins.—The question of the origin of the paired fins is much more difficult and is still far from settled, although many, perhaps the majority of recent writers favor the theory that these fins are parts of a once continuous lateral fold of skin, corresponding to the vertical fold which forms the dorsal, anal, and caudal. In this view the lateral fold, at first continuous, became soon atrophied in the middle, while at either end it is highly specialized, at first into an organ of direction, then into fan-shaped and later paddle-shaped organs of locomotion. According to another view, the paired fins originated from gill structures, originally both close behind the head, the ventral fin migrating backward with the progress of evolution of the species.
Evidence of Paleontology.—If we had representations of all the early forms of fishes arranged in proper sequence, we could decide once for all, by evidence of paleontology, which form of fin appears first and what is the order of appearance. As to this, it is plain that we do not know the most primitive form of fin. Sharks of unknown character must have existed long before the earliest remains accessible to us. Hence the evidence of paleontology seems conflicting and uncertain. On the whole it lends most support to the fin-fold theory. In the later Devonian, a shark, Cladoselache fyleri, is found in which the paired fins are lappet-shaped, so formed and placed as to suggest their origin from a continuous fold of skin. In this species the dorsal fins show much the same form. Other early sharks, constituting the order of Acanthodei, have fins somewhat similar, but each preceded by a stiff spine, which may be formed from coalescent rays.

Fig. 51.—Cladoselache fyleri (Newberry), restored. Upper Devonian of Ohio. (After Dean.)

Fig. 52.—Fold-like pectoral and ventral fins of Cladoselache fyleri. (After Dean.)
Long after these appears another type of sharks represented by Pleuracanthus and Cladodus, in which the pectoral fin is a jointed organ fringed with rays arranged serially in one or two rows. This form of fin has no resemblance to a fold of skin, but accords better with Gegenbaur's theory that the pectoral limb was at first a modified gill-arch. In the Coal Measures are found also teeth of sharks (Orodontidæ) which bear a strong resemblance to still existing forms of the family of Heterodontidæ, which originates in the Permian. The existing Heterodontidæ have the usual specialized form of shark-fin, with three of the basal segments especially enlarged and placed side by side, the type seen in modern sharks. Whatever the primitive form of shark-fin, it may well be doubted whether any one of these three (Cladoselache, Pleuracanthus, or Heterodontus) actually represents it. The beginning is therefore unknown, though there is some evidence that Cladoselache is actually more nearly primitive than any of the others. As we shall see, the evidence of comparative anatomy may be consistent with either of the two chief theories, while that of ontogeny or embryology is apparently inconclusive, and that of paleontology is apparently most easily reconciled with the theory of the fin-fold.
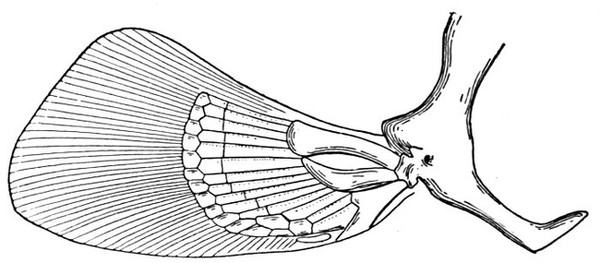
Fig. 53.—Pectoral fin of shark, Chiloscyllium. (After Parker and Haswell.)
Development of the Paired Fins in the Embryo.—According to Dr. John A. Ryder ("Embryography of Osseous Fishes," 1882) "the paired fins in Teleostei arise locally, as short longitudinal folds, with perhaps a few exceptions. The pectorals of Lepisosteus originate in the same way. Of the paired fins, the pectoral or anterior pair seems to be the first to be developed, the ventral or pelvic pair often not making its appearance until after the absorption of the yolk-sac has been completed, in other cases before that event, as in Salmo and in Gambusia. The pectoral fin undergoes less alteration of position during its evolution than the posterior pair."
In the codfish (Gadus callarias) the pectoral fin-fold "appears as a slight longitudinal elevation of the skin on either side of the body of the embryo a little way behind the auditory vesicles, and shortly after the tail of the embryo begins to bud out. At the very first it appears to be merely a dermal fold, and in some forms a layer of cells extends out underneath it from the sides of the body, but does not ascend into it. It begins to develop as a very low fold, hardly noticeable, and, as growth proceeds, its base does not expand antero-posteriorly, but tends rather to become narrowed, so that it has a pedunculated form. With the progress of this process the margin of the fin-fold also becomes thinner at its distal border, and at the basal part mesodermal cells make their appearance more noticeably within the inner contour-line. The free border of the fin-fold grows out laterally and longitudinally, expanding the portion outside of the inner contour-line of the fin into a fan-shape. This distal thinner portion is at first without any evidence of rays; further than that there is a manifest tendency to a radial disposition of the histological elements of the fin."
The next point of interest is found in the change of position of the pectoral fin by a rotation on its base. This is associated with changes in the development of the fish itself. The ventral fin is also, in most fishes, a short horizontal fold and just above the preanal part of the median vertical fold which becomes anal, caudal, and dorsal. But in the top-minnow (Gambusia), of the order Haplomi, the ventral first appears as "a little papilla and not as a fold, where the body-walls join the hinder upper portion of the yolk-sac, a very little way in front of the vent." "These two modes of origin," observes Dr. Ryder, "are therefore in striking contrast and well calculated to impress us with the protean character of the means at the disposal of Nature to achieve one and the same end."
Current Theories as to Origin of Paired Fins.—There are three chief theories as to the morphology and origin of the paired fins. The earliest is that of Dr. Karl Gegenbaur, supported by various workers among his students and colleagues. In his view the pectoral and ventral fins are derived from modifications of primitive gill-arches. According to this theory, the skeletal arrangements of the vertebrate limb are derived from modifications of one primitive form, a structure made up of successive joints, with a series of fin-rays on one or both sides of it. To this structure Gegenbaur gives the name of archipterygium. It is found in the shark, Pleuracanthus, in Cladodus, and in all the Dipnoan and Crossopterygian fishes, its primitive form being still retained in the Australian genus of Dipnoans, Neoceratodus. This biserial archipterygium with its limb-girdle is derived from a series of gill-rays attached to a branchial arch. The backward position of the ventral fin is due to a succession of migrations in the individual and in the species.
As to this theory, Mr. J. Graham Kerr observes:
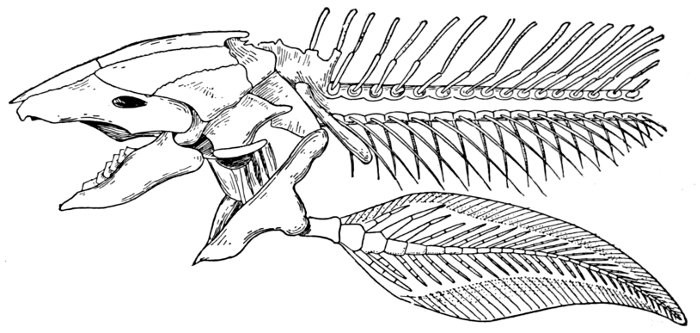
Fig. 54.—Skull and shoulder-girdle of Neoceratodus forsteri (Günther), showing the archipterygium.
"The Gegenbaur theory of the morphology of vertebrate limbs thus consists of two very distinct portions. The first, that the archipterygium is the ground-form from which all other forms of presently existing fin skeletons are derived, concerns us only indirectly, as we are dealing here only with the origin of the limbs, i.e., their origin from other structures that were not limbs.
"It is the second part of the view that we have to do with, that deriving the archipterygium, the skeleton of the primitive paired fin, from a series of gill-rays and involving the idea that the limb itself is derived from the septum between two gill-clefts.
"This view is based on the skeletal structures within the fin. It rests upon (1) the assumption that the archipterygium is the primitive type of fin, and (2) the fact that amongst the Selachians is found a tendency for one branchial ray to become larger than the others, and, when this has happened, for the base of attachment of neighboring rays to show a tendency to migrate from the branchial arch on to the base of the larger or, as we may call it, primary ray; a condition coming about which, were the process to continue rather farther than it is known to do in actual fact, would obviously result in a structure practically identical with the archipterygium. Gegenbaur suggests that the archipterygium actually has arisen in this way in phylogeny."

Fig. 55.—Acanthoessus wardi (Egerton). Carboniferous. Family Acanthoessidæ. (After Woodward.)

Fig. 56.—Shoulder-girdle of Acanthoessus. (After Dean.)
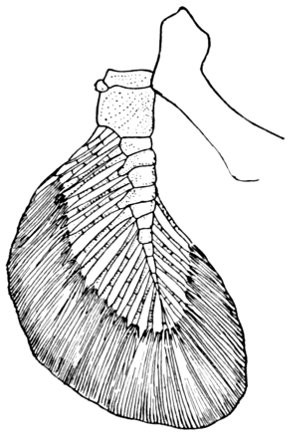
Fig. 57.—Pectoral fin of Pleuracanthus. (After Dean.)
The fin-fold theory of Balfour, adopted by Dohrn, Weidersheim, Thacher, Mivart, Ryder, Dean, Boulenger, and others, and now generally accepted by most morphologists as plausible, is this: that "The paired limbs are persisting and exaggerated portions of a fin-fold once continuous, which stretched along each side of the body and to which they bear an exactly similar phylogenetic relation as do the separate dorsal and anal fins to the once continuous median fin-fold."
"This view, in its modern form, was based by Balfour on his observation that in the embryos of certain Elasmobranchs the rudiments of the pectoral and pelvic fins are at a very early period connected together by a longitudinal ridge of thickened epiblast—of which indeed they are but exaggerations. In Balfour's own words referring to these observations: 'If the account just given of the development of the limb is an accurate record of what really takes place, it is not possible to deny that some light is thrown by it upon the first origin of the vertebrate limbs. The facts can only bear one interpretation, viz., that the limbs are the remnants of continuous lateral fins.'
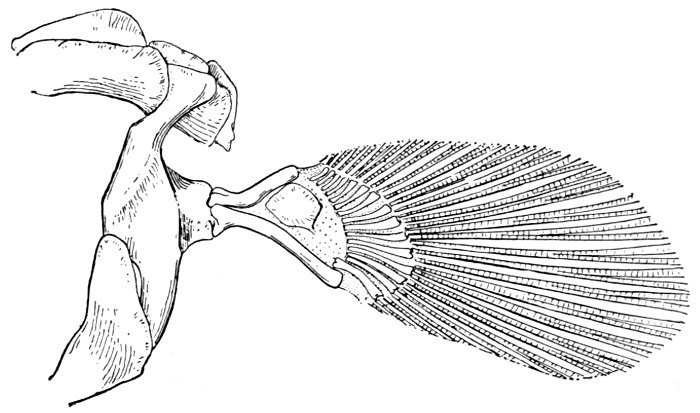
Fig. 58.—Shoulder-girdle of Polypterus bichir. Specimen from the White Nile.
"A similar view to that of Balfour was enunciated almost synchronously by Thacher and a little later by Mivart—in each case based on anatomical investigation of Selachians—mainly relating to the remarkable similarity of the skeletal arrangements in the paired and unpaired fins."
A third theory is suggested by Mr. J. Graham Kerr (Cambridge Philos. Trans., 1899), who has recently given a summary of the theories on this subject. Mr. Kerr agrees with Gegenbaur as to the primitive nature of the archipterygium, but believes that it is derived, not from the gill-septum, but from an external gill. Such a gill is well developed in the young of all the living sharks, Dipnoans and Crossopterygians, and in the latter types of fishes it has a form analogous to that of the archipterygium, although without bony or cartilaginous axis.
We may now take up the evidence in regard to each of the different theories, using in part the language of Kerr, the paragraphs in quotation-marks being taken from his paper. We may first consider Balfour's theory of the lateral fold.
Balfour's Theory of the Lateral Fold.—"The evidence in regard to this view may be classed under three heads, as ontogenetic, comparative anatomical, and paleontological. The ultimate fact on which it was founded was Balfour's discovery that in certain Elasmobranch embryos, but especially in Torpedo (Narcobatus), the fin rudiments were, at an early stage, connected by a ridge of epiblast. I am not able to make out what were the other forms in which Balfour found this ridge, but subsequent research, in particular by Mollier, a supporter of the lateral-fold view, is to the effect that it does not occur in such ordinary sharks as Pristiurus and Mustelus, while it is to be gathered from Balfour himself that it does not occur in Scyllium (Scyliorhinus).
"It appears to me that the knowledge we have now that the longitudinal ridge is confined to the rays and absent in the less highly specialized sharks greatly diminishes its security as a basis on which to rest a theory. In the rays, in correlation with their peculiar mode of life, the paired fins have undergone (in secondary development) enormous extension along the sides of the body, and their continuity in the embryo may well be a mere foreshadowing of this.
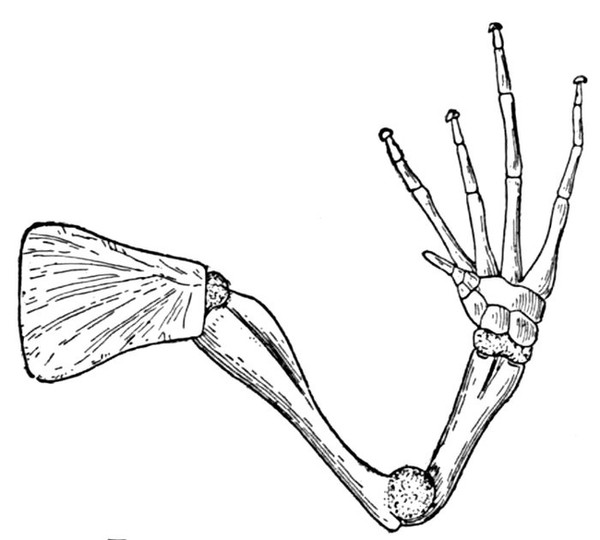
Fig. 59.—Arm of a frog.
"An apparently powerful support from the side of embryology came in Dohrn and Rabl's discoveries that in Pristiurus all the interpterygial myotomes produce muscle-buds. This, however, was explained away by the Gegenbaur school as being merely evidence of the backward migration of the hind limb—successive myotomes being taken up and left behind again as the limb moved farther back. As either explanation seems an adequate one, I do not think we can lay stress upon this body of facts as supporting either one view or the other. The facts of the development of the skeleton cannot be said to support the fold view; according to it we should expect to find a series of metameric supporting rays produced which later on become fused at their bases. Instead of this we find a longitudinal bar of cartilage developing quite continuously, the rays forming as projections from its outer side.
"The most important evidence for the fold view from the side of comparative anatomy is afforded by (1) the fact that the limb derives its nerve supply from a large number of spinal nerves, and (2) the extraordinary resemblance met with between the skeletal arrangements of paired and unpaired fins. The believers in the branchial arch hypothesis have disposed of the first of these in the same way as they did the occurrence of interpterygial myotomes, by looking on the nerves received from regions of the spinal cord anterior to the attachment of the limb as forming a kind of trail marking the backward migration of the limb.
"The similarity in the skeleton is indeed most striking, though its weight as evidence has been recently greatly diminished by the knowledge that the apparently metameric segmentation of the skeletal and muscular tissues of the paired fins is quite secondary and does not at all agree with the metamery of the trunk. What resemblance there is may well be of a homoplastic character when we take into account the similarity in function of the median and unpaired fins, especially in such forms as Raja, where the anatomical resemblances are especially striking. There is a surprising dearth of paleontological evidence in favor of this view."
The objection to the first view is its precarious foundation. Such lateral folds are found only in certain rays, in which they may be developed as a secondary modification in connection with the peculiar form of these fishes. Professor Kerr observes that this theory must be looked upon and judged: "Just as any other view at the present time regarding the nature of the vertebrate limb, rather as a speculation, brilliant and suggestive though it be, than as a logically constructed theory of the now known facts. It is, I think, on this account allowable to apply to it a test of a character which is admittedly very apt to mislead, that of 'common sense.'
"If there is any soundness in zoological speculation at all, I think it must be admitted that the more primitive vertebrates were creatures possessing a notochordal axial skeleton near the dorsal side, with the main nervous axis above it, the main viscera below it, and the great mass of muscle lying in myotomes along its sides. Now such a creature is well adapted to movements of the character of lateral flexure, and not at all for movements in the sagittal plane—which would be not only difficult to achieve, but would tend to alternately compress and extend its spinal cord and its viscera. Such a creature would swim through the water as does a Cyclostome, or a Lepidosiren, or any other elongated vertebrate without special swimming organs. Swimming like this, specialization for more and more rapid movement would mean flattening of the tail region and is extension into an at first not separately mobile median tail-fold. It is extremely difficult to my mind to suppose that a new purely swimming arrangement should have arisen involving up-and-down movement, and which, at its first beginnings, while useless as a swimming organ itself, must greatly detract from the efficiency of that which already existed."
Objections to Gegenbaur's Theory.—We now return to the Gegenbaur view—that the limb is a modified gill-septum.
"Resting on Gegenbaur's discovery already mentioned, that the gill-rays in certain cases assume an arrangement showing great similarity to that of the skeletal elements of the archipterygium, it has, so far as I am aware, up to the present time received no direct support whatever of a nature comparable with that found for the rival view in the fact that, in certain forms at all events, the limbs actually do arise in the individual in the way that the theory holds they did in phylogeny. No one has produced either a form in which a gill-septum becomes the limb during ontogeny, or the fossil remains of any form which shows an intermediate condition.
"The portion of Gegenbaur's view which asserts that the biserial archipterygial fin is of an extremely primitive character is supported by a large body of anatomical facts, and is rendered further probable by the great frequency with which fins apparently of this character occur amongst the oldest known fishes. On the lateral-fold view we should have to regard these as independently evolved, which would imply that fins of this type are of a very perfect character, and in that case we may be indeed surprised at their so complete disappearance in the more highly developed forms, which followed later on."
Fig. 60.—Pleuracanthus decheni (Goldfuss). (After Dean.)
As to Gegenbaur's theory it is urged that no form is known in which a gill-septum develops into a limb during the growth of the individual. The main thesis, according to Professor Kerr, "that the archipterygium was derived from gill-rays, is supported only by evidence of an indirect character. Gegenbaur in his very first suggestion of his theory pointed out, as a great difficulty in the way of its acceptance, the position of the limbs, especially of the pelvic limbs, in a position far removed from that of the branchial arches. This difficulty has been entirely removed by the brilliant work of Gegenbaur's followers, who have shown from the facts of comparative anatomy and embryology that the limbs, and the hind limbs especially, actually have undergone, and in ontogeny do undergo, an extensive backward migration. In some cases Braus has been able to find traces of this migration as far forward as a point just behind the branchial arches. Now, when we consider the numbers, the enthusiasm, and the ability of Gegenbaur's disciples, we cannot help being struck by the fact that the only evidence in favor of this derivation of the limbs has been that which tends to show that a migration of the limbs backwards has taken place from a region somewhere near the last branchial arch, and that they have failed utterly to discover any intermediate steps between gill-rays and archipterygial fin. And if for a moment we apply the test of common sense we cannot but be impressed by the improbability of the evolution of a gill-septum, which in all the lower forms of fishes is fixed firmly in the body-wall, and beneath its surface, into an organ of locomotion.

Fig. 61.—Embryos of Heterodontus japonicus Maclay and Macleay, a Cestraciont shark, showing the backward migration of the gill-arches and the forward movement of the pectoral fin. a, b, c, representing different stages of growth. (After Dean.)
"May I express the hope that what I have said is sufficient to show in what a state of uncertainty our views are regarding the morphological nature of the paired fins, and upon what an exceedingly slender basis rest both of the two views which at present hold the field?"
As to the backward migration of the ventral fins, Dr. Bashford Dean has recently brought forward evidence from the embryo of a very ancient type of shark (Heterodontus japonicus) that this does not actually occur in that species. On the other hand, we have a forward migration of the pectoral fin, which gradually takes its place in advance of the hindmost gill-arches. The accompanying cut is from Dean's paper, "Biometric Evidence in the Problem of the Paired Limbs of the Vertebrates" (American Naturalist for November, 1902). Dean concludes that in Heterodontus "there is no evidence that there has ever been a migration of the fins in the Gegenbaurian sense." "The gill region, at least in its outer part, shows no affinity during proportional growth with the neighboring region of the pectoral fin. In fact from an early stage onward, they are evidently growing in opposite directions."
Kerr's Theory of Modified External Gills.—"It is because I feel that in the present state of our knowledge neither of the two views I have mentioned has a claim to any higher rank than that of extremely suggestive speculations that I venture to say a few words for the third view, which is avowedly a mere speculation.
"Before proceeding with it I should say that I assume the serial homology of fore and hind limbs to be beyond dispute. The great and deep-seated resemblances between them are such as to my mind seem not to be adequately explicable except on this assumption.
"In the Urodela (salamanders) the external gills are well-known structures—serially arranged projections from the body-wall near the upper ends of certain of the branchial arches. When one considers the ontogenetic development of these organs, from knob-like outgrowth from the outer face of the branchial arch, covered with ectoderm and possessing a mesoblastic core, and which frequently if not always appear before the branchial clefts are open, one cannot but conclude that they are morphologically projections of the outer skin and that they have nothing whatever to do with the gill-pouches of the gut-wall. Amongst the Urodela one such gill projects from each of the first three branchial arches. In Lepidosiren there is one on each of the branchial arches I-IV. In Polypterus and Calamoichthys (Erpetoichthys) there is one on the hyoid arch. Finally, in many Urodelan larvæ we have present at the same time as the external gills a pair of curious structures called balancers. At an early stage of my work on Lepidosiren, while looking over other vertebrate embryos and larvæ for purposes of comparison, my attention was arrested by these structures, and further examinations, by section or otherwise, convinced me that there were serial homologues of the external gills, situated on the mandibular arch. On then looking up the literature, I found that I was by no means first in this view. Rusconi had long ago noticed the resemblance, and in more recent times both Orr and Maurer had been led to the same conclusion as I had been. Three different observers having been independently led to exactly the same conclusions, we may, I think, fairly enough regard the view I have mentioned of the morphological nature of the balancers as probably a correct one.
"Here, then, we have a series of homologous structures projecting from each of the series of visceral arches. They crop up on the Crossopterygii, the Dipnoi, and the Urodela, i.e., in three of the most archaic of the groups of Gnathostomata. But we may put it in another way. The groups in which they do not occur are those whose young possess a very large yolk-sac (or which are admittedly derived from such forms). Now wherever we have a large yolk-sac we have developed on its surface a rich network of blood-vessels for purposes of nutrition. But such a network must necessarily act as an extraordinarily efficient organ of respiration, and did we not know the facts we might venture to prophesy that in forms possessing it any other small skin-organ of respiration would tend to disappear.
"No doubt these external gills are absent also in a few of the admittedly primitive forms such as, e.g., (Neo-) Ceratodus. But I would ask that in this connection one should bear in mind one of the marked characteristics of external gills—their great regenerative power. This involves their being extremely liable to injury and consequently a source of danger to their possessor. Their absence, therefore, in certain cases may well have been due to natural selection. On the other hand, the presence in so many lowly forms of these organs, the general close similarity in structure that runs through them in different forms, and the exact correspondence in their position and relations to the body can, it seems to me, only be adequately explained by looking on them as being homologous structures inherited from a common ancestor and consequently of great antiquity in the vertebrate stem."
As to the third theory, Professor Kerr suggests tentatively that the external gill may be the structure modified to form the paired limbs. Of the homology of fore and hind limbs and consequently of their like origin there can be no doubt.
The general gill-structures have, according to Kerr, "the primary function of respiration. They are also, however, provided with an elaborate muscular apparatus comprising elevators, depressors, and adductors, and larvæ possessing them may be seen every now and then to give them a sharp backward twitch. They are thus potentially motor organs. In such a Urodele as Amblystoma their homologues on the mandibular arch are used as supporting structures against a solid substratum exactly as are the limbs of the young Lepidosiren.
[3] Philos. Trans., Lond., 1900.
In the genus Lepidosiren, also a Dipnoan, the pectoral limb has the same axial structure, but is without fin-rays, although in the breeding season the posterior limb or ventral fin in the male is covered with a brush of fine filaments. This structure, according to Prof. J. G. Kerr,[3] is probably without definite function, but belongs to the "category of modifications so often associated with the breeding season (cf. the newts' crest) commonly called ornamental, but which are perhaps more plausibly looked upon as expressions of the intense vital activity of the organisms correlated with its period of reproductive activity." Professor Kerr, however, thinks it not unlikely that this brush of filaments with its rich blood-supply may serve in the function of respiration, a suggestion first made by Professor Lankester.

Fig. 62.—Polypterus congicus, a Crossopterygian fish from the Congo River. Young, with external gills. (After Boulenger.)
"I have, therefore, to suggest that the more ancient Gnathostomata possessed a series of potentially motor, potentially supporting structures projecting from their visceral arches; it was inherently extremely probable that these should be made use of when actual supporting, and motor appendages had to be developed in connection with clambering about a solid substratum. If this had been so, we should look upon the limb as a modified external gill; the limb-girdle, with Gegenbaur, as a modified branchial arch.
"This theory of the vertebrate paired limb seems to me, I confess, to be a more plausible one on the face of it than either of the two which at present hold the field. If untrue, it is so dangerously plausible as to surely deserve more consideration than it appears to have had. One of the main differences between it and the other two hypotheses is that, instead of deriving the swimming-fin from the walking and supporting limb, it goes the other way about. That this is the safer line to take seems to me to be shown by the consideration that a very small and rudimentary limb could only be of use if provided with a fixed point d'appui. Also on this view, the pentadactyle limb and the swimming-fin would probably be evolved independently from a simple form of limb. This would evade the great difficulties which have beset those who have endeavored to establish the homologies of the elements of the pentadactyle limb with those of any type of fully formed fin."
Uncertain Conclusions.—In conclusion we may say that the evidence of embryology in this matter is inadequate, though possibly favoring on the whole the fin-fold theory; that of morphology is inconclusive, and probably the final answer may be given by paleontology. If the records of the rocks were complete, they would be decisive. At present we have to decide which is the more primitive of two forms of pectoral fin actually known among fossils. That of Cladoselache is a low, horizontal fold of skin, with feeble rays, called by Cope ptychopterygium. That of Pleuracanthus is a jointed paddle-shaped appendage with a fringe of rays on either side. In the theory of Gegenbaur and Kerr Pleuracanthus must be, so far as the limbs are concerned, the form nearest the primitive limb-bearing vertebrate. In Balfour's theory Cladoselache is nearest the primitive type from which the other and with it the archipterygium of later forms may be derived.
Boulenger and others question even this, believing that the archipterygium in Pleuracanthus and other primitive sharks and that in Neoceratodus and its Dipnoan and Crossopterygian allies and ancestors have been derived independently, not the latter from the former. In this view there is no real homology between the archipterygium in the sharks possessing it and that in the Dipnoans and Crossopterygians. In the one theory the type of Pleuracanthus would be ancestral to the other sharks on the one hand, and to Crossopterygians and all higher vertebrates on the other. With the theory of the origin of the pectoral from a lateral fold, Pleuracanthus would be merely a curious specialized offshoot from the primitive sharks, without descendants and without special significance in phylogeny.
As elements bearing on this decision we may note that the tapering unspecialized diphycercal tail of Pleuracanthus seems very primitive in comparison with the short heterocercal tail of Cladoselache. This evidence, perhaps deceptive, is balanced by the presence on the head of Pleuracanthus of a highly specialized serrated spine, evidence of a far from primitive structure. Certainly neither the one genus nor the other actually represents the primitive shark. But as Cladoselache appears in geological time, long before Pleuracanthus, Cladodus, or any other shark with a jointed, archipterygial fin, the burden of proof, according to Dean, rests with the followers of Gegenbaur. If the remains found in the Ordovician at Cañon City referred to Crossopterygians are correctly interpreted, we must regard the shark ancestry as lost in pre-Silurian darkness, for in sharks of some sort the Crossopterygians apparently must find their remote ancestry.
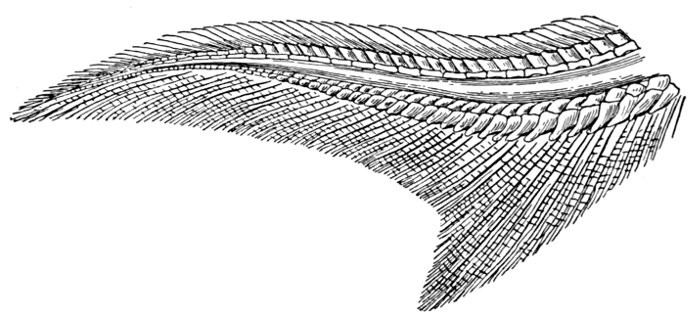
Fig. 63.—Heterocercal tail of Sturgeon, Acipenser sturio (Linnæus). (After Zittel.)
Forms of the Tail in Fishes.—In the process of development the median or vertical fins are, as above stated, older than the paired fins or limbs, whatever be the origin of the latter. They arise in a dermal keel, its membranes fitting and accentuating the undulatory motion of the body.
In this elementary fin-fold slender supports (actinotrichia), the rudiments of fin-rays, appear at intervals. In those fins of most service in the movement of the fish, the fin-rays are strengthened, and their basal supports specialized.
Dean calls attention to the fact that in fishes which swim, when adult, by an undulatory motion, the paired fins tend to disappear, as in the eel and in all eel-like fishes, as blennies and eel-pouts.
The form of the tail at the base of the caudal fin varies in the different groups. In most primitive types, as in most embryonic fishes, the vertebræ grow smaller to the last (diphycercal). In others, also primitive, the end of the tail is directed upward, and the most of the caudal fin is below it. Such a tail is seen in most sharks, in the sturgeon, garpike, bowfin, and in the Ganoid fishes. It is known as heterocercal, and finally in ordinary fishes the tail becomes homocercal or fan-shaped, although usually some trace of the heterocercal condition is traceable, gradually growing less with the process of development.
Since Professor Agassiz first recognized, in 1833, the distinction between the heterocercal and homocercal tail, this matter has been the subject of elaborate investigation and a number of additional terms have been proposed, some of which are in common use.
A detailed discussion of these is found in a paper by Dr. John A. Ryder "On the Origin of Heterocercy" in the Report of the U. S. Fish Commissioner for 1884. In this paper a dynamic or mechanical theory of the causes of change of form is set forth, parts of this having a hypothetical and somewhat uncertain basis.
Dr. Ryder proposes the name archicercal to denote the cylindroidal worm-like caudal end of the larva of fishes and amphibians before they acquire median fin-folds. The term lophocercal is proposed by Ryder for the form of caudal fin which consists of a rayless fold of skin continuous with the skin of the tail, the inner surfaces of this fold being more or less nearly in contact. To the same type of tail Dr. Jeffries Wyman in 1864 gave the name protocercal. This name was used for the tail of the larval ray when it acquires median fin-folds. The term implies, what cannot be far from true, that this form of tail is the first in the stages of evolution of the caudal fin.
To the same type of tail Mr. Alexander Agassiz gave, in 1877, the name of leptocardial, on the supposition that it represented the adult condition of the lancelet. In this creature, however, rudimentary basal rays are present, a condition differing from that of the early embryos.
The diphycercal tail, as usually understood, is one in which the end of the vertebral column bears "not only hypural but also epural intermediary pieces which support rays." The term is used for the primitive type of tail in which the vertebræ, lying horizontally, grow progressively smaller, as in Neoceratodus, Protopterus, and other Dipnoans and Crossopterygians. The term was first applied by McCoy to the tails of the Dipnoan genera Diplopterus and Gyroptychius, and for tails of this type it should be reserved.
Fig. 64.—Heterocercal tail of Bowfin, Amia calva (Linnæus). (After Zittel.)

Fig. 65.—Heterocercal tail of Garpike, Lepisosteus osseus (Linnæus).
The heterocercal tail is one in which the hindmost vertebræ are bent upwards. The term is generally applied to those fishes only in which this bending is considerable and is externally evident, as in the sharks and Ganoids. The character disappears by degrees, changing sometimes to diphycercal or leptocercal by a process of degeneration, or in ordinary fishes becoming homocercal. Dr. Ryder uses the term heterocercal for all cases in which any up-bending of the axis takes place, even though it involves the modification of but a single vertebra. With this definition, the tail of salmon, herring, and even of most bony fishes would be considered heterocercal, and most or all of these pass through a heterocercal stage in the course of development. The term is, however, usually restricted to those forms in which the curving of the axis is evident without dissection.
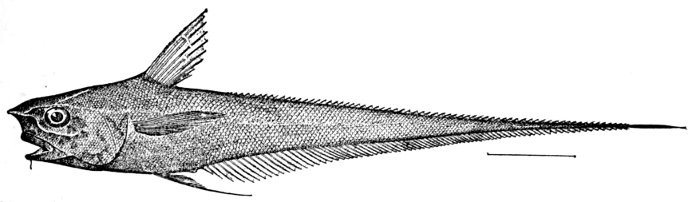
Fig. 66.—Coryphænoides carapinus (Goode and Bean), showing leptocercal tail. Gulf Stream.
The homocercal tail is the fan-shaped or symmetrical tail common among the Teleosts, or bony fishes. In its process of development the individual tail is first archicercal, then lophocercal, then diphycercal, then heterocercal, and lastly homocercal. A similar order is indicated by the sequence of fossil fishes in the rocks, although some forms of diphycercal tail may be produced by degeneration of the heterocercal tail, as suggested by Dr. Dollo and Dr. Boulenger, who divide diphycercal tails into primitive and secondary.
The peculiar tapering tail of the cod, the vertebræ growing progressively smaller behind, is termed isocercal by Professor Cope. This form differs little from diphycercal, except in its supposed derivation from the homocercal type. A similar form is seen in eels.
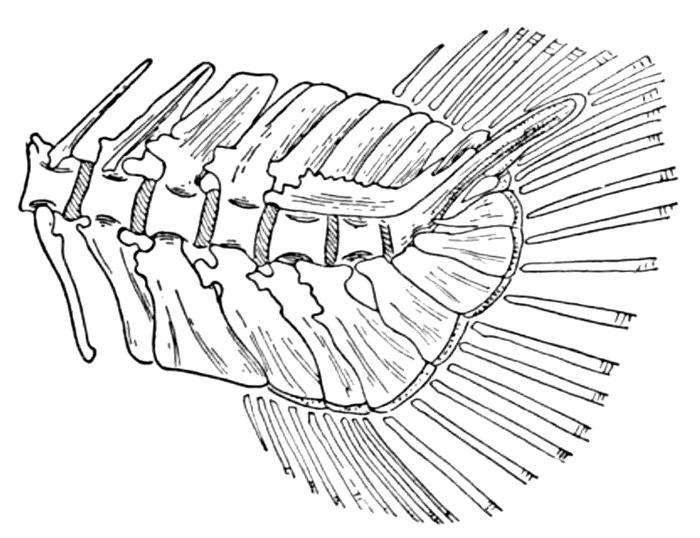
Fig. 67.—Heterocercal tail of Young Trout, Salmo fario (Linnæus). (After Parker and Haswell.)
The term leptocercal has been suggested by Gaudry, 1883, for those tails in which the vertebral column ends in a point. We may, perhaps, use it for all such as are attenuate, ending in a long point or whip, as in the Macrouridæ, or grenadiers, the sting-rays, and in various degenerate members of almost every large group.
The term gephyrocercal is devised by Ryder for fishes in which the end of the vertebral axis is aborted in the adult, leaving the caudal elements to be inserted on the end of this axis, thus bridging over the interval between the vertical fins, as the name (γεφύρος, bridge; κέρκος, tail) is intended to indicate. Such a tail has been recognized in four genera only, Mola, Ranzania, Fierasfer, and Echiodon, the head-fishes and the pearl-fishes.
Fig. 68.—Isocercal tail of Hake, Merluccius productus (Ayres).
Fig. 69.—Homocercal tail of a Flounder, Paralichthys californicus.
The part of the body of the fish which lies behind the vent is known as the urosome. The urostyle is the name given to a modified bony structure, originally the end of the notochord, turned upward in most fishes. The term opisthure is suggested by Ryder for the exserted tip of the vertebral column, which in some larvæ (Lepisosteus) and in some adult fishes (Fistularia, Chimæra) projects beyond the caudal fin. The urosome, or posterior part of the body, must be regarded as a product of evolution and specialization, its function being largely that of locomotion. In the theoretically primitive fish there is no urosome, the alimentary canal, as in the worm, beginning at one end of the body and terminating at the other.

Fig. 70.—Gephyrocercal tail of Mola mola (Linnæus). (After Ryder.)
Homologies of the Pectoral Limb.—Dr. Gill has made an elaborate attempt to work out the homologies of the bones of the pectoral limb.[4] From his thesis we take the following:
"The following are assumed as premises that will be granted by all zootomists:
"1. Homologies of parts are best determinable, ceteris paribus, in the most nearly related forms.
"2. Identification should proceed from a central or determinate point outwards.
"The applications of these principles are embodied in the following conclusions:
"1. The forms that are best comparable and that are most nearly related to each other are the Dipnoi, an order of fishes at present represented by Lepidosiren, Protopterus, and Ceratodus, and the Batrachians as represented by the Ganocephala, Salamanders, and Salamander-like animals.
"2. The articulation of the anterior member with the shoulder-girdle forms the most obvious and determinable point for comparison in the representatives of the respective classes.
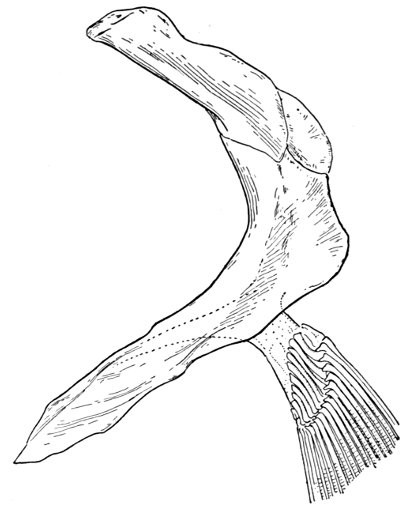
Fig. 71.—Shoulder-girdle of Amia calva (Linnæus).
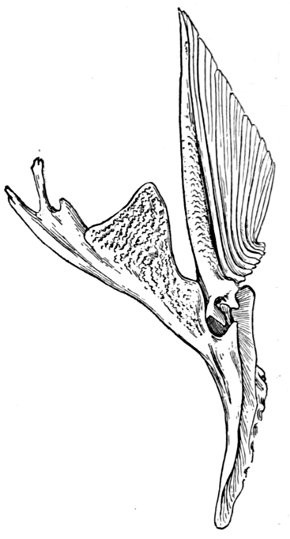
Fig. 72.—Shoulder-girdle of a Sea Catfish, Selenaspis dowi.
The Girdle in Dipnoans.—"The proximal element of the anterior limb in the Dipnoi has almost by common consent been regarded as homologous with the humerus of the higher vertebrates.
"The humerus of Urodele Batrachians, as well as the extinct Ganocephala and Labyrinthodontia, is articulated chiefly with the coracoid. Therefore the element of the shoulder-girdle with which the humerus of the Dipnoi is articulated must also be regarded as the coracoid (subject to the proviso hereinafter stated), unless some specific evidence can be shown to the contrary. No such evidence has been produced.
"The scapula in the Urodele and other Batrachians is entirely or almost wholly excluded from the glenoid foramen, and above the coracoid. Therefore the corresponding element in Dipnoi must be the scapula.
"The other elements must be determined by their relation to the preceding, or to those parts from or in connection with which they originate. All those elements in immediate connection with the pectoral fin and the scapula must be homologous as a whole with the coraco-scapular plate of the Batrachians; that is, it is infinitely more probable that they represent, as a whole or as dismemberments therefrom, the coraco-scapular element than that they independently originated. But the homogeneity of that coraco-scapular element forbids the identification of the several elements of the fish's shoulder-girdle with regions of the Batrachian's coraco-scapular plate.

Fig. 73.—Clavicles of a Sea Catfish, Selenaspis dowi (Gill).
"And it is equally impossible to identify the fish's elements with those of the higher reptiles or other vertebrates which have developed from the Batrachians. The elements in the shoulder-girdles of the distantly separated classes may be (to use the terms introduced by Dr. Lankester) homoplastic, but they are not homogenetic. Therefore they must be named accordingly. The element of the Dipnoan's shoulder-girdle, continuous downward from the scapula, and to which the coracoid is closely applied, may be named ectocoracoid.
"Neither the scapula in Batrachians nor the cartilaginous extension thereof, designated suprascapula, is dissevered from the coracoid. Therefore there is an a priori improbability against the homology with the scapula of any part having a distant and merely ligamentous connection with the humerus-bearing element. Consequently, as an element better representing the scapula exists, the element named scapula (by Owen, Günther, etc.) cannot be the homologue of the scapula of Batrachians. On the other hand, its more intimate relations with the skull and the mode of development indicate that it is rather an element originating and developed in more intimate connection with the skull. It may therefore be considered, with Parker, as a post-temporal.
"The shoulder-girdle in the Dipnoi is connected by an azygous differentiated cartilage, swollen backwards. It is more probable that this is the homologue of the sternum of Batrachians, and that in the latter that element has been still more differentiated and specialized than that it should have originated de novo from an independently developed nucleus."
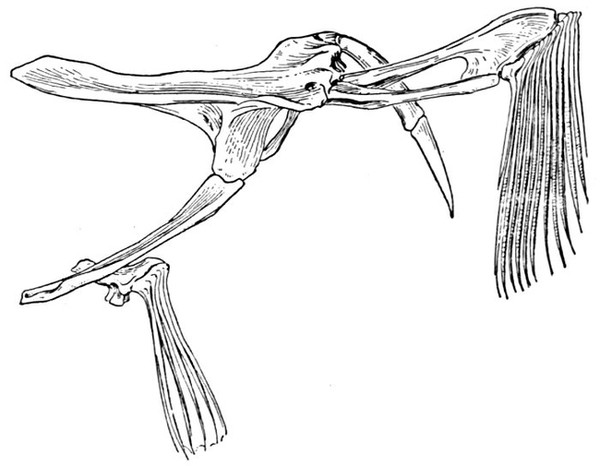
Fig. 74.—Shoulder-girdle of a Batfish, Ogcocephalus radiatus (Mitchill).
The Girdle in Fishes Other than Dipnoans.—"Proceeding from the basis now obtained, a comparative examination of other types of fishes successively removed by their affinities from the Lepidosirenids may be instituted.
"With the humerus of the Dipnoans, the element of the Polypterids (single at the base, but immediately divaricating and with its limbs bordering an intervening cartilage which supports the pectoral and its basilar ossicles) must be homologous. But it is evident that the external elements of the so-called carpus of the teleosteoid Ganoids are homologous with that element in Polypterids. Therefore those elements cannot be carpal, but must represent the humerus.
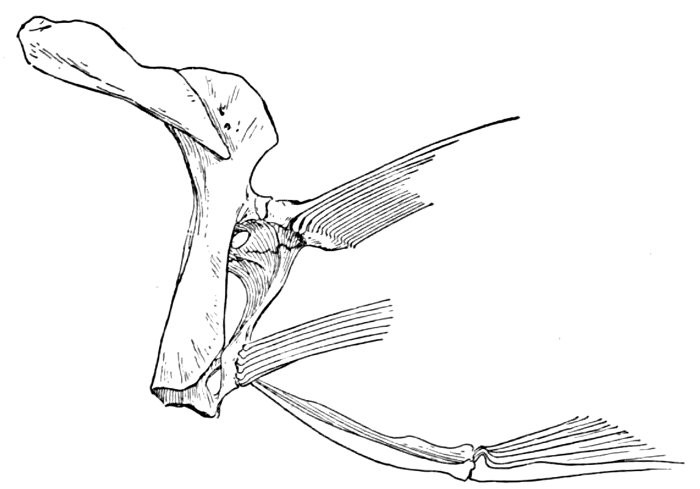
Fig. 75.—Shoulder-girdle of a Threadfin, Polydactylus approximans (Lay and Bennett).
"The element with which the homologue of the humerus, in Polypterids, is articulated must be homologous with the analogous element in Dipnoans, and therefore with the coracoid. The coracoid of Polypterids is also evidently homologous with the corresponding element in the other Ganoids, and the latter consequently must be also coracoid. It is equally evident, after a detailed comparison, that the single coracoid element of the Ganoids represents the three elements developed in the generalized Teleosts (Cyprinids, etc.) in connection with the basis of the pectoral fin, and, such being the case, the nomenclature should correspond. Therefore the upper element may be named hypercoracoid; the lower, hypocoracoid; and the transverse or median, mesocoracoid.
"The two elements of the arch named by Parker, in Lepidosiren, 'supraclavicle' (scapula) and 'clavicle' (ectocoracoid) seem to be comparable together, and as a whole, with the single element carrying the humerus and pectoral fin in the Crossopterygians (Polypterus and Calamoichthys) and other fishes, and therefore not identical respectively with the 'supraclavicle' and 'clavicle' (except in part) recognized by him in other fishes. As this compound bone, composed of the scapula and ectocoracoid fused together, has received no name which is not ambiguous or deceptive in its homologous allusions, it may be designated as proscapula.
"The post-temporal of the Dipnoans is evidently represented by the analogous element in the Ganoids generally, as well as in the typical fishes. The succeeding elements (outside those already alluded to) appear from their relations to be developed from or in connection with the post-temporal, and not from the true scapular apparatus; they may therefore be named post-temporal, posterotemporal, and teleo-temporal. It will be thus seen that the determinations here adopted depend mainly (1) on the interpretation of the homologies of the elements with which the pectoral limbs are articulated, and (2) on the application of the term 'coracoid.' The name 'coracoid,' originally applied to the process so called in the human scapula and subsequently extended to the independent element homologous with it in birds and other vertebrates, has been more especially retained (e.g., by Parker in mammals, etc.) for the region including the glenoid cavity. On the assumption that this may be preferred by some zootomists, the preceding terms have been applied. But if the name should be restricted to the proximal element, nearest the glenoid cavity, in which ossification commences, the name paraglenal given by Dugès to the cartilaginous glenoid region can be adopted, and the coracoid would then be represented (in part) rather by the element so named by Owen. That eminent anatomist, however, reached his conclusion (only in part the same as that here adopted) by an entirely different course of reasoning, and by a process, as it may be called, of elimination; that is, recognizing first the so-called 'radius' and 'ulna,' the 'humerus,' the 'scapula,' and the 'coracoid' were successively identified from their relations to the elements thus determined and because they were numerically similar to the homonymous parts among higher vertebrates."
FOOTNOTES:
[4] Catalogue of the Families of Fishes, 1872.
CHAPTER VI
THE ORGANS OF RESPIRATION

How Fishes Breathe.—The fish breathes the air which is dissolved in water. It cannot use the oxygen which is a component part of water, nor can it, as a rule, make use of atmospheric air. The amount of oxygen required for the low vegetative processes of the fish is comparatively small. According to Dr. Günther, a man consumes 50,000 times as much oxygen as a tench. But some fishes demand more oxygen than others. Some, like the catfish or the loach, will survive long out of water, while others die almost instantly if removed from their element or if the water is allowed to become foul. In most cases the temperature of the blood of the fish is but little above that of the water in which they live, but in the mackerel and other muscular fishes the temperature of the body may be somewhat higher.
Some fishes which live in mud, especially in places which become dry in summer, have special contrivances by which they can make use of atmospheric air. In a few primitive fishes (Dipnoans, Crossopterygians, Ganoids) the air-bladder retains its original function of a lung. In other cases some peculiar structure exists in connection with the gills. Such a contrivance for holding water above the gills is seen in the climbing perch of India (Anabas scandens) and other members of the group called Labyrinthici.
In respiration, in fishes generally, the water is swallowed through the mouth and allowed to pass out through the gill-openings, thus bathing the gills. In a few of the lower types a breathing-pore takes the place of the gill-openings.
The gills, or branchiæ, are primarily folds of the skin lining the branchial cavity. In most fishes they form fleshy fringes or laminæ throughout which the capillaries are distributed. In the embryos of sharks, skates, chimæras, lung-fishes, and Crossopterygians external gills are developed, but in the more specialized forms these do not appear outside the gill-cavity. In some of the sharks, and especially the rays, a spiracle or open foramen remains behind the eye. Through this spiracle, leading from the outside into the cavity of the mouth, water is drawn downwards to pass outward over the gills. The presence of this breathing-hole permits these animals to lie on the bottom without danger of inhaling sand.

Fig. 76.—Gill-basket of Lamprey. (After Dean.)
The Gill-structures.—The three main types of gills among fishes are the following: (a) the purse-shaped gills found in the hagfishes and lampreys, known as a class as Marsipobranchs, or purse-gills. These have a number (5 to 12) of sac-like depressions on the side of the body, lined with gill-fringes and capillaries, the whole supported by an elaborate branchial basket formed of cartilage. (b) The plate-gills, found among the sharks, rays, and chimæras, thence called Elasmobranchs, or plate-gills. In these the gill-structures are flat laminæ, attached by one side to the gill-arches. (c) The fringe-gills found in ordinary fishes, in which the gill-filaments containing the capillaries are attached in two rows to the outer edge of each gill-arch. The so-called tuft-gills (Lophobranchs) of the sea-horse and pipefish are like these in structure, but the filaments are long, while the arches are very short. In most of the higher fishes a small accessory gill (pseudobranchia) is developed in the skin of the inner side of the opercle.
The Air-bladder.—The air-bladder, or swim-bladder, must be classed among the organs of respiration, although in the higher fishes its functions in this regard are rudimentary, and in some cases it has taken collateral functions (as a hydrostatic organ of equilibrium, or perhaps as an organ of hearing) which have no relation to its original purpose.
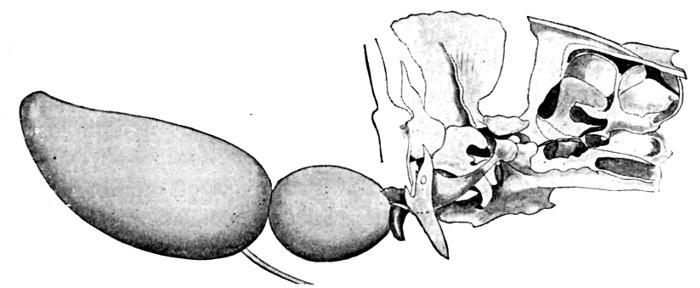
Fig. 77.—Weberian apparatus and air-bladder of Carp. (From Günther, after Weber.)
The air-bladder is an internal sac possessed by many fishes, but not by all. It lies in the dorsal part of the abdominal cavity above the intestines and below the kidneys. In some cases it is closely adherent to the surrounding tissues. In others it is almost entirely free, lying almost loose in the cavity of the body. In some cases it is enclosed in a bony capsule. In the allies of the carp and catfish, which form the majority of fresh-water fishes, its anterior end is connected through a chain of modified vertebræ to the ear. Sometimes its posterior end fits into an enlarged and hollow interhæmal bone. Sometimes, again, a mass of muscle lies in front of it or is otherwise attached to it. Sometimes it is divided into two or three parts by crosswise constrictions. Sometimes it is constricted longitudinally, and at other times it has attached to it a complication of supplemental tubes of the same character as the air-bladder itself. In still other cases it is divided by many internal partitions into a cellular body, similar to the lung of the higher vertebrates, though the cells are coarser and less intricate. This condition is evidently more primitive than that of the empty sac.
The homology of the air-bladder with the lung is evident. This is often expressed in the phrase that the lung is a developed air-bladder. This is by no means true. To say that the air-bladder is a modified and degenerate lung is much nearer the truth, although we should express the fact more exactly to say that both air-bladder and lung are developed from a primitive cellular breathing-sac, originally a diverticulum from the ventral walls of the œsophagus.
The air-bladder varies in size as much as in form. In some fishes it extends from the head to the tail, while in others it is so minute as to be scarcely traceable. It often varies greatly in closely related species. The common mackerel (Scomber scombrus) has no air-bladder, while in the closely related colias or chub mackerel (Scomber japonicus) the organ is very evident. In other families, as the rockfishes (Scorpænidæ), genera with and those without the air-bladder are scarcely distinguishable externally. In general, fishes which lie on the bottom, those which inhabit great depths, and those which swim freely in the open sea, as sharks and mackerel, lack the air-bladder. In the sharks, rays, and chimæras there is no trace of an air-bladder. In the mackerel and other bony fishes without it, it is lost in the process of development.
The air-bladder is composed of two layers of membrane, the outer one shining, silvery in color, with muscular fibres, the inner well supplied by blood-vessels. The gas within the air-bladder must be in most cases secreted from the blood-vessels. In river fishes it is said to be nearly pure nitrogen. In marine fishes it is mostly oxygen, with from 6 to 10 per cent of carbonic-acid gas, while in the deep-sea fishes oxygen is greatly in excess. In Lopholatilus, a deep-sea fish, Professor R. W. Tower finds 66 to 69 per cent of oxygen. In Trigla lyra Biot records 87 per cent. In Dentex dentex, a shore fish of Europe, 40 per cent of oxygen was found in the air-bladder. Fifty per cent is recorded from the European porgy, Pagrus pagrus. In a fish dying from suffocation the amount of carbonic-acid gas (CO2) is greatly increased, amounting, according to recent researches of Professor Tower on the weak-fish, Cynoscion regalis, to 24 to 29 per cent. This shows conclusively that the air-bladder is to some degree a reservoir of oxygen secreted from the blood, to which channel it may return through a kind of respiration.
The other functions of the air-bladder have been subject to much question and are still far from understood. The following summary of the various views in this regard we copy from Professor Tower's paper on "The Gas in the Swim-bladder of Fishes":
"The function of the swim-bladder of fishes has attracted the attention of scientists for many centuries. The rôle that this structure plays in the life of the animal has been interpreted in almost as many ways as there have been investigators, and even now there is apparently much doubt as to the true functions of the swim-bladder. Consequently any additional data concerning this organ are of immediate scientific value.
"Aristotle, writing about the noises made by fishes, states that 'some produce it by rubbing the gill-arches ...; others by means of the air-bladder. Each of these fishes contains air, by rubbing and moving of which the noise is produced.' The bladder is thus considered a sound-producing organ, and it is probable that he arrived at this result by his own investigations.
"Borelli (De Motu Animalium, 1680) attributed to the air-bladder a hydrostatic function which enabled the fish to rise and fall in the water by simply distending or compressing the air-bladder. This hypothesis, which gives to the fish a volitional control over the air-bladder—it being able to compress or distend the bladder at pleasure—has prevailed, to a greater or less degree, from the time of Borelli to the present. To my knowledge, however, there are no investigations which warrant such a theory, while, on the other hand, there are many facts, as shown by Moreau's experiment, which distinctly contradict this belief. Delaroche (Annales du Mus. d'Hist. Nat., tome XIV, 1807-1809) decidedly opposed the ideas of Borelli, and yet advanced an hypothesis similar to it in many respects. Like Borelli, he said that the fish could compress or dilate the bladder by means of certain muscles, but this was to enable the fish to keep the same specific gravity as the surrounding medium, and thus be able to remain at any desired depth (and not to rise or sink). This was also disproved later by Moreau. Delaroche proved that there existed a constant exchange between the air in the air-bladder and the air in the blood, although he did not consider the swim-bladder an organ of respiration.
"Biot (1807), Provençal and Humboldt (1809), and others made chemical analyses of the gas in the swim-bladder, and found 1 to 5 per cent of CO2, 1 to 87 per cent of O2, and the remainder nitrogen. The most remarkable fact discovered about this mixture was that it frequently consisted almost entirely of oxygen, the per cent of oxygen increasing with the depth of the water inhabited by the fish. The reasons for this phenomenon have never been satisfactorily explained.
"In 1820 Weber described a series of paired ossicles which he erroneously called stapes, malleus, and incus, and which connected the air-bladder in certain fishes with a part of the ear—the atrium sinus imparis. Weber considered the swim-bladder to be an organ by which sounds striking the body from the outside are intensified, and these sounds are then transmitted to the ear by means of the ossicles. The entire apparatus would thus function as an organ of hearing. Weber's views remained practically uncontested for half a century, but recently much has been written both for and against this theory. Whatever the virtues of the case may be, there is certainly an inviting field for further physiological investigations regarding this subject, and more especially on the phenomena of hearing in fishes.
"Twenty years later Johannes Müller described, in certain Siluroid fishes, a mechanism, the so-called 'elastic-spring' apparatus, attached to the anterior portion of the air-bladder, which served to aid the fish in rising and sinking in the water according as the muscles of this apparatus were relaxed or contracted to a greater or lesser degree. This interpretation of the function of the 'elastic-spring' mechanism was shown by Sörensen to be untenable. Müller also stated that in some fish, at least, there was an exchange of gas between blood and air-bladder—the latter having a respiratory function—and regarded the gas in the air-bladder as the result of active secretion. In Malapterurus (Torpedo electricus) he stated that it is a sound-producing organ.
"Hasse, in 1873, published the results of his investigations on the functions of the ossicles of Weber, stating that their action was that of a manometer, acquainting the animal with the degree of pressure that is exerted by the gases in the air-bladder against its walls. This pressure necessarily varies with the different depths of water which the fish occupies. Hasse did not agree with Weber that the ear is affected by the movements of these ossicles.
"One year later Dufosse described in some fishes an air-bladder provided with extrinsic muscles by whose vibration sound was produced, the sound being intensified by the air-bladder, which acted as a resonator. He also believed that certain species produced a noise by forcing the gas from the air-bladder through a pneumatic duct.
"At about the same time Moreau published his classical work on the functions of the air-bladder. He proved by ingenious experiments that many of the prevailing ideas about the action of the air-bladder were erroneous, and that this organ serves to equilibrate the body of the fish with the water at any level. This is not accomplished quickly, but only after sufficient time for the air in the bladder to become adjusted to the increase or decrease in external pressure that has taken place. The fish, therefore, makes no use of any muscles in regulating the volume of its air-bladder. The animal can accommodate itself only gradually to considerable changes in depth of water, but can live equally comfortably at different depths, provided that the change has been gradual enough. Moreau's experiments also convinced him that the gas is actually secreted into the air-bladder, and that there is a constant exchange of gas between it and the blood. In these investigations he has also noticed that section of the sympathetic-nerve fibres supplying the walls of the air-bladder hastens the secreting of the gas into the empty bladder. Since then Bohr has shown that section of the vagus nerve causes the secretion to cease. Moreau noticed in one fish (Trigla) having an air-bladder supplied with muscles that the latter served to make the air-bladder produce sound.
"Again, in 1885, the Weberian mechanism was brought to our attention with a new function attributed to it by Sagemehl who stated that this mechanism exists not for any auditory purposes, nor to tell the fish at what level of the water it is swimming, but to indicate to the fish the variations in the atmospheric pressure. Sörensen tersely contrasts the views of Hasse and Sagemehl by saying that 'Hasse considers the air-bladder with the Weberian mechanism as a manometer; Sagemehl regards it as a barometer.' The theory of Sagemehl has, naturally enough, met with little favor. Sörensen (1895) held that there is but little evidence for attributing to the air-bladder the function of a lung. It is to be remembered, however, that, according to Sörensen's criterion no matter what exchange of gases takes place between blood and air-bladder, it cannot be considered an organ of respiration, 'unless its air is renewed by mechanical respiration.'
"Sörensen also refutes, from anatomical and experimental grounds, the many objections to Weber's theory of the function of the ossicles. He would thus attribute to the air-bladder the function of hearing; indeed in certain species the only reason for the survival of the air-bladder is that 'the organ is still of acoustic importance; that it acts as a resonator.' This idea, Sörensen states, is borne out by the anatomical structure found in Misgurnus and Chlarias, which resembles the celebrated 'Colladon resonator.' This author attributes to the air-bladder with its 'elastic spring' and various muscular mechanisms the production of sound as its chief function."
Origin of the Air-bladder.—In the more primitive forms, and probably in the embryos of all species, the air-bladder is joined to the œsophagus by an air-duct. This duct is lost entirely in the adult of all or nearly all of the thoracic and jugular fishes, and in some of the abdominal forms. The lancelets, lampreys, sharks, rays, and chimæras have no air-bladder, but in the most primitive forms of true fishes (Dipnoans and Crossopterygians), having the air-bladder cellular or lung-like, the duct is well developed, freely admitting the external air which the fish may rise to the surface to swallow. In most fishes the duct opens into the œsophagus from the dorsal side, but in the more primitive forms it enters from the ventral side, like the windpipe of the higher vertebrates. In some of the Dipnoans the air-bladder divides into two parts, in further resemblance to the true lungs.
The Origin of the Lungs.—The following account of the function of the air-bladder and of its development and decline is condensed from an article by Mr. Charles Morris:[5]
"If now we seek to discover the original purpose of this organ, there is abundant reason to believe that it had nothing to do with swimming. Certainly the great family of the sharks, which have no bladder, are at no disadvantage in changing their depth or position in the water. Yet if the bladder is necessary to any fish as an aid in swimming, why not to all? And if this were its primary purpose, how shall we explain its remarkable variability? No animal organ with a function of essential importance presents such extraordinary modifications in related species and genera. In the heart, brain, and other organs there is one shape, position, and condition of greatest efficiency, and throughout the lower forms we find a steady advance towards this condition. Great variation, on the other hand, usually indicates that the organ is of little functional importance, or that it has lost its original function. Such we conceive to be the case with the air-bladder. The fact of its absence from some and its presence in other fishes of closely related species goes far to prove that it is a degenerating organ; and the same is shown by the fact that it is useless in some species for the purpose to which it is applied in others. That it had, at some time in the past, a function of essential importance there can be no question. That it exists at all is proof of this. But its modern variations strongly indicate that it has lost this function and is on the road towards extinction. Larval conditions show that it had originally a pneumatic duct as one of its essential parts, but this has in most cases disappeared. The bladder itself has in many cases partly or wholly disappeared. Where preserved, it seems to be through its utility for some secondary purpose, such as an aid in swimming or in hearing. That its evolution began very long ago there can be no question; and the indications are that it began long ago to degenerate, through the loss of its primitive function.
"What was this primitive function? In attempting to answer this question we must first consider the air-bladder in relation to the fish tribe as a whole. No shark or ray possesses the air-bladder. In some few sharks, indeed, there is a diverticulum of the pharynx which may be a rudimentary approach to the air-bladder; but this is very questionable. The conditions of its occurrence in the main body of modern fishes, the Teleostean, we have already considered. But in the most ancient living orders of fishes it exists in an interesting condition. In every modern Dipnoan, Crossopterygian, and Ganoid the air-bladder has an effective pneumatic duct. This in the Ganoids opens into the dorsal side of the œsophagus, but in the Dipnoans and Crossopterygians, like the windpipe of lung-breathers, it opens into the ventral side. In the Dipnoans, also survivors from the remote past, the duct not only opens ventrally into the œsophagus, but the air-bladder does duty as a lung. Externally it differs in no particular from an air-bladder; but internally it presents a cellular structure which nearly approaches that of the lung of the batrachians. There are three existing representatives of the Dipnoans. One of these, the Australian lung-fish (Neoceratodus) has a single bladder, which, however, is provided with breathing-pouches having a symmetrical lateral arrangement. It has no pulmonary artery, but receives branches from the arteria cœliaca. In the other two forms, Lepidosiren and Protopterus, the kindred 'mudfishes' of the Amazon basin and tropical Africa, the bladder or lung is divided into two lateral chambers, as in the land animals, and is provided with a separate pulmonary artery.
"The opinion seems to have been tacitly entertained by physiologists that this employment of the air-bladder by the Dipnoans as a lung is a secondary adaptation, a side issue from its original purpose. It is more likely that this is the original purpose, and that its degeneration is due to the disappearance of the necessity of such a function. As regards the gravitative employment of the bladder, the Teleostean fishes, to which this function is confined, are of comparatively modern origin; while the Dipnoans are surviving representatives of a very ancient order of fishes, which flourished in the Devonian age of geology, and in all probability breathed air then as now; and the Crossopterygians and Ganoids, which approach them in this particular, are similarly ancient in origin, and were the ancestors of the Teleosteans. The natural presumption, therefore, is that the duty which it subserved in the most ancient fishes was its primitive function.
"The facts of embryology lend strong support to this hypothesis. For the air-bladder is found to arise in a manner very similar to the development of the lung. They each begin as an outgrowth from the fore part of the alimentary tract, the only difference being that the air-bladder usually rises dorsally and the lung ventrally. The fact already cited, that the pneumatic duct is always present in the larval form in fishes that possess a bladder, is equally significant. All the facts go to show that the introduction of external air into the body was a former function of the air-bladder, and that the atrophy of the duct in many cases, and the disappearance of the bladder in others, are results of the loss of this function.
"Such an elaborate arrangement for the introduction of air into the body could have, if we may judge from analogy, but one purpose, that of breathing, to which purpose the muscular and other apparatus for compressing and dilating the bladder, now seemingly adapted to gravitative uses, may have been originally applied. The same may be said of the great development of blood-capillaries in the inner tunic of the bladder. These may now be used only for the secretion of gas into its interior, but were perhaps originally employed in the respiratory secretion of oxygen. In fact all the circumstances mentioned—the similarity in larval development between the bladder and lung, the larval existence of the pneumatic duct, the arrangements for compressing and dilating the bladder, and the capillary vessels on its inner tunic—point to the breathing of air as its original purpose.
"It is probable that the Ganoid, as well as the Dipnoan, air-bladder is to some extent still used in breathing. The Dipnoans have both lungs and gills, and probably breathe with the latter in ordinary cases, but use their lungs when the inland waters in which they live become thick and muddy, or are charged with gases from decomposing organic matter. The Ganoid fishes to some extent breathe the air. In Polypterus the air-bladder resembles the Dipnoan lung in having lateral divisions and a ventral connection with the œsophagus, while in Lepisosteus (the American garpike) it is cellular and lung-like. This fish keeps near the surface, and may be seen to emit air-bubbles, probably taking in a fresh supply of air. The American bowfin, or mudfish (Amia), has a bladder of the same lung-like character, and has been seen to come to the surface, open its jaws widely, and apparently swallow a large quantity of air. He considers that both Lepisosteus and Amia inhale and exhale air at somewhat regular intervals, resembling in this the salamanders and tadpoles, 'which, as the gills shrink and the lungs increase, come more frequently to the surface for air.'
"As the facts stand there is no evident line of demarcation between the gas-containing bladders of many of the Teleosteans, the air-containing bladders of the others and the Ganoids, and the lung of the Dipnoans, and the indications are in favor of their having originally had the same function, and of this being the breathing of air.
"If now we ask what were the conditions of life under which this organ was developed, and what the later conditions which rendered it of no utility as a lung, some definite answer may be given. The question takes us back to the Devonian and Silurian geological periods, during which the original development of the bladder probably took place. In this era the seas were thronged with fishes of several classes, the Elasmobranchs among others, followed by the Dipnoi and Crossopterygians. The sharks were without, the Dipnoans and Crossopterygians doubtless with, an air-bladder—a difference in organization which was most likely due to some marked difference in their life-habits. The Elasmobranchs were the monarchs of the seas, against whose incursions the others put on a thick protective armor, and probably sought the shallow shore waters, while their foes held chief possession of the deeper waters without.
"We seem, then, to perceive the lung-bearing fishes, driven by their foes into bays and estuaries, and the waters of shallow coasts, ascending streams and dwelling in inland waters. Here two influences probably acted on them. The waters they dwelt in were often thick with sediment, and were doubtless in many instances poorly aerated, rendering gill-breathing difficult. And the land presented conditions likely to serve as a strong inducement to fishes to venture on shore. Its plant-life was abundant, while its only animal inhabitants seem to have been insects, worms, and snails. There can be little doubt that the active fish forms of that period, having no enemies to fear on the land, and much to gain, made active efforts to obtain a share of this vegetable and animal food. Even to-day, when they have numerous foes to fear, many fishes seek food on the shore, and some even climb trees for this purpose. Under the conditions of the period mentioned there was a powerful inducement for them to assume this habit.
"Such conditions must have strongly tended to induce fishes to breathe the air, and have acted to develop an organ for this purpose. In addition to the influences of foul or muddy water and of visits to land may be named that of the drying-out of pools, by which fishes are sometimes left in the moist mud till the recurrence of rains, or are even buried in the dried mud during the rainless season. This is the case with the modern Dipnoi, which use their lungs under such circumstances. In certain other fresh-water fishes, of the family Ophiocephalidæ, air is breathed while the mud continues soft enough for the fish to come to the surface, but during the dry period the animal remains in a torpid state. These fishes have no lungs, but breathe the air into a simple cavity in the pharynx, whose opening is partly closed by a fold of the mucous membrane. Other Labyrinthici, of similar habits, possess a more developed breathing organ. This is a cavity formed by the walls of the pharynx, in which are thin laminæ, or plates, which undoubtedly perform an oxygenating function. The most interesting member of this family is Anabas scandens, the climbing perch. In this fish, which not only leaves the water, but is said to climb trees, the air-breathing organ is greatly developed. The labyrinthici, moreover, have usually large air-bladders. As regards the occasional breathing of air by fishes, even in species which do not leave the water, it is quite common, particularly among fresh-water species. Cuvier remarks that air is perhaps necessary to every kind of fish; and that, particularly when the atmosphere is warm, most of our lacustrine species sport on the surface for no other purpose.
"It is not difficult to draw a hypothetical plan of the development of the air-bladder as a breathing organ. In the two families of fishes just mentioned, whose air-bladders indicate that they once possessed the air-breathing function and have lost it, we perceive the process of formation of an air-breathing organ beginning over again under stress of similar circumstances. The larval development of the air-bladder points significantly in the same direction. In fact we have strong reason to believe that air-breathing in fishes was originally performed, as it probably often is now, by the unchanged walls of the œsophagus. Then these walls expanded inwardly, forming a simple cavity, partly closed by a fold of membrane, like that of the Ophiocephalidæ. A step further reduced this membranous fold to a narrow opening, leading to an inner pouch. As the air-breathing function developed, the opening became a tube, and the pouch a simple lung, with compressing muscles and capillary vessels. By a continuation of the process the smooth-walled pouch became sacculated, its surface being increased by folding into breathing cells. Finally, a longitudinal constriction divided it into two lateral pouches, such as we find in the lung of the Dipnoans. This brings us to the verge of the lung of the amphibians, which is but a step in advance, and from that the line of progress is unbroken to the more intricate lung of the higher land animals.
"The dorsal position of the bladder and its duct would be a difficulty in this inquiry, but for the fact that the duct is occasionally ventral. This dorsal position may have arisen from the upward pressure of air in the swimming fish, which would tend to lift the original pouch. But in the case of fishes which made frequent visits to the shore new influences must have come into play. The effect of gravity tended to draw the organ and its duct downward, as we find in the Crossopterygians and in all the Dipnoans, and its increased use in breathing required a more extended surface. Through this requirement came the pouched and cellular lung of the Dipnoans. Of every stage of the process here outlined examples exist, and there is great reason to believe that the development of the lung followed the path above pointed out.
"When the carboniferous era opened there may have been many lung- and gill-breathing fishes which spent much of their time on land, and some of which, by a gradual improvement of their organs of locomotion, changed into batrachians. But with the appearance of the latter, and of their successors, the reptiles, the relations of the fish to the land radically changed. The fin, or the simple locomotor organ, of the Dipnoans could not compete with the leg and foot as organs of land locomotion, and the fish tribe ceased to be lords of the land, where, instead of feeble prey, they now found powerful foes, and were driven back to their native habitat, the water. Nor did the change end here. In time the waters were invaded by the reptiles, numerous swimming forms appearing, which it is likely were abundant in the shallower shore-line of the ocean, while they sent many representatives far out to sea. These were actively carnivorous, making the fish their prey, the great mass of whom were doubtless driven into the deeper waters, beyond the reach of their air-breathing foes.
"In this change of conditions we seem to perceive an adequate cause for the loss of air-breathing habits in those fishes in which the lung development had not far progressed. It may indeed have been a leading influence in the development of the Teleostean or bony fishes, as it doubtless was in the loss of its primitive function by, and the subsequent changes of, the air-bladder.
"Such of the Crossopterygians and Dipnoans as survived in their old condition had to contend with adverse circumstances. Most of them in time vanished, while their descendants which still exist have lost in great measure their air-breathing powers, and the Dipnoans, in which the development of the lung had gone too far for reversal, have degenerated into eel-like, mud-haunting creatures, in which the organs of locomotion have become converted into the feeble paddle-like limbs of Neoceratodus and the filamentary appendages of the other species.
"As regards the presence of a large quantity of oxygen in the bladders of deep-swimming marine fishes, it not unlikely has a respiratory purpose, the bladder being, as suggested by Semper, used as a reservoir for oxygen, to serve the fish when sleeping, or when, from any cause, not actively breathing. The excess of oxygen is not due to any like excess in the gaseous contents of sea-water, for the percentage of oxygen decreases from the surface downward, while that of nitrogen remains nearly unchanged. In all cases, indeed, the bladder may preserve a share of its old function, and act as an aid in respiration. Speaking of this, Cuvier says: 'With regard to the presumed assistance which the swim-bladder affords in respiration, it is a fact that when a fish is deprived of that organ, the production of carbonic acid by the branchiæ is very trifling,' thus strongly indicating that the bladder still plays a part in the oxygenation of the blood.
"Under the hypothesis here presented the process of evolution involved may be thus summed up. Air-breathing in fishes was originally performed by the unchanged walls of the œsophagus perhaps at specially vascular localities. Then the wall folded inward, and a pouch was finally formed, opening to the air. The pouch next became constricted off, with a duct of connection. Then the pouch became an air-bladder with respiratory function, and finally developed into a simple lung. These air-breathing fishes haunted the shores, their fins becoming converted into limbs suitable for land locomotion, and in time developed into the lung- and gill-breathing batrachia, and these in their turn into the lung-breathing reptilia, the locomotor organs gradually increasing in efficiency. Of these pre-batrachia we have existing representatives in the mud-haunting Dipnoi, with their feeble limbs. In the great majority of the Ganoid fishes the bladder served but a minor purpose as a breathing organ, the gills doing the bulk of the work. In the Teleostean descendants of the Ganoids the respiratory function of the bladder in great measure or wholly ceased, in the majority of cases the duct closing up or disappearing, leaving the pouch as a closed internal sac, far removed from its place of origin. In this condition it served as an aid in swimming, perhaps as a survival of one of its ancient uses. It gained also in certain cases some connection with the organ of hearing. But these were makeshift and unimportant functions, as we may gather from the fact that many fishes found no need for them, the bladder, in these cases, decreasing in size until too small to be of use in swimming, and in other cases completely disappearing after having travelled far from its point of origin. In some other cases, above cited, the process seems to have begun again, in modern times, in an eversion of the wall of the œsophagus for respiratory purposes. The whole process, if I have correctly conceived it, certainly forms a remarkable organic cycle of development and degeneration, which perhaps has no counterpart of similarly striking character in the whole range of organic life."
The Heart of the Fish.—The heart of the fish is simple in structure, small in size, and usually placed far forward, just behind the branchial cavity, and separated from the abdominal cavity by a sort of "diaphragm" formed of thickened peritoneum. In certain eels the heart is remote from the head.
The heart consists of four parts, the sinus venosus, into which the veins enter, the auricle or atrium, the ventricle, and the arterial bulb at the base of the great artery which carries the blood to the gills. Of these parts the ventricle is deepest in color and with thickest walls. The arterial bulb varies greatly in structure, being in the sharks, rays, Ganoids, and Dipnoans muscular and provided with a large number of internal valves, and contracting rhythmically like the ventricle. In the higher fishes these structures are lost, the walls of the arterial bulb are not contractile, and the interior is without valves, except the pair that separate it from the ventricle.
In the lancelet there is no proper heart, the function of the heart being taken by a contractile blood-vessel situated on the ventral side of the alimentary canal. In the Dipnoans, which are allied to the ancestors of the higher vertebrates, there is the beginning of a division of the ventricle, and sometimes of the auricle, into parts by a median septum. In the higher vertebrates this septum becomes more and more specialized, separating auricle and ventricle into right and left cavities. The blood in the fish is not returned to the heart after purification, but is sent directly over the body.
The Flow of Blood.—The blood in fishes is thin and pale red (colorless in the lancelet) and with elliptical blood-corpuscles. It enters the sinus venosus from the head through the jugular vein, from the kidney and body walls through the cardinal vein, and from the liver through the hepatic veins. Hence it passes to the auricle and ventricle, and from the ventricle through the arterial bulb, or conus arteriosus to the ventral aorta. Thence it flows to the gills, where it is purified. After passing through the capillaries of the gill-filaments it is collected in paired arteries from each pair of gills. These vessels unite to form the dorsal aorta, which extends the length of the body just below the back-bone. From the dorsal aorta the subclavian arteries branch off toward the pectoral fins. From a point farther back arise the mesenteric arteries carrying blood to the stomach, intestine, liver, and spleen. In the tail the caudal vein carries blood to the kidneys. These secrete impurities arising from waste of tissues, after which the blood again passes to the heart through the cardinal vein. From the intestine the blood, charged with nutritive materials in solution, is carried by the portal vein to the liver. Here it again passes by the hepatic sinus to the sinus venosus and the heart.
The details of the circulatory system vary a good deal in the different groups, and a comparative study of the direction of veins and arteries is instructive and interesting.
The movement of the blood in fishes is relatively slow, and its temperature is raised but little above that of the surrounding water.
[4] Catalogue of the Families of Fishes, 1872.
Homologies of the Pectoral Limb.—Dr. Gill has made an elaborate attempt to work out the homologies of the bones of the pectoral limb.[4] From his thesis we take the following:
[5] The Origin of Lungs: A Chapter in Evolution. American Naturalist, December, 1892.
FOOTNOTES:
[5] The Origin of Lungs: A Chapter in Evolution. American Naturalist, December, 1892.
CHAPTER VII
THE NERVOUS SYSTEM

The nerves of the Fish.—The nervous system in the fish, as in the higher vertebrates, consists of brain and spinal cord with sensory, or afferent, and motor, or efferent, nerves. As in other vertebrates, the nerve substance is divided into gray matter and white matter, or nerve-cells and nerve-fibres. In the fish, however, the whole nervous system is relatively small, and the gray matter less developed than in the higher forms. According to Günther the brain in the pike (Esox) forms but 1/1305 part of the weight of the body; in the burbot (Lota) about 1/720 part.
The cranium in fishes is relatively small, but the brain does not nearly fill its cavity, the space between the dura mater, which lines the skull-cavity, and the arachnoid membrane, which envelops the brain, being filled with a soft fluid containing a quantity of fat.
The Brain of the Fish.—It is most convenient to examine the fish-brain, first in its higher stages of development, as seen in the sunfish, striped bass, or perch. As seen from above the brain of a typical fish seems to consist of five lobes, four of them in pairs, the fifth posterior to these and placed on the median line. The posterior lobe is the cerebellum, or metencephalon, and it rests on the medulla oblongata, the posterior portion of the brain, which is directly continuous with the spinal cord.
In front of the cerebellum lies the largest pair of lobes, each of them hollow, the optic nerves being attached to the lower surface. These are known as the optic lobes, or mesencephalon. In front of these lie the two lobes of the cerebrum, also called the hemispheres, or prosencephalon. These lobes are usually smaller than the optic lobes and solid. In some fishes they are crossed by a furrow, but are never corrugated as in the brain of the higher animals. In front of the cerebrum lie the two small olfactory lobes, which receive the large olfactory nerve from the nostrils. From its lower surface is suspended the hypophysis or pituitary gland.
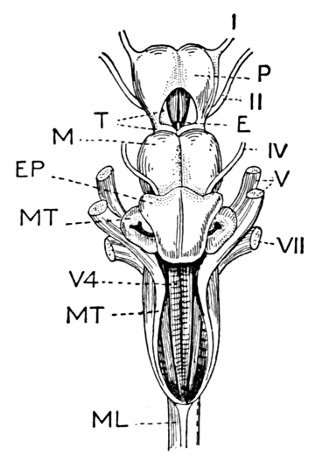
Fig. 78.—Brain of a Shark (Squatina squatina L.). (After Dean.)
- I. First cranial nerve (olfactory).
- P. Prosencephalon (cerebrum).
- E. Epiphysis.
- T. Thalamencephalon.
- II. Second cranial nerve.
- IV. Fourth cranial nerve.
- V. Fifth cranial nerve.
- VII. Seventh cranial nerve.
- V4. Fourth ventricle.
- M. Mesencephalon (optic lobes).
- MT. Metencephalon (medulla).
- EP. Epencephalon (cerebellum).
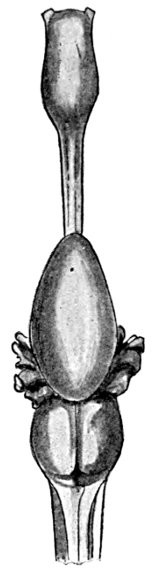
Fig. 79.—Brain of Chimæra monstrosa. (After Wilder per Dean.)
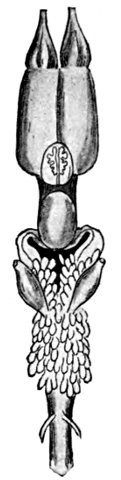
Fig. 80.—Brain of Protopterus annectens. (After Burckhardt per Dean.)
In most of the bony fishes the structure of the brain does not differ materially from that seen in the perch. In the sturgeon, however, the parts are more widely separated. In the Dipnoans the cerebral hemispheres are united, while the optic lobe and cerebellum are very small. In the sharks and rays the large cerebral hemispheres are usually coalescent into one, and the olfactory nerves dilate into large ganglia below the nostrils. The optic lobes are smaller than the hemispheres and also coalescent. The cerebellum is very large, and the surface of the medulla oblongata is more or less modified or specialized. The brain of the shark is relatively more highly developed than that of the bony fishes, although in most other regards the latter are more distinctly specialized.
The Pineal Organ.—Besides the structures noted in other fishes the epiphysis, or pineal organ, is largely developed in sharks, and traces of it are found in most or all of the higher vertebrates. In some of the lizards this epiphysis is largely developed, bearing at its tip a rudimentary eye. This leaves no doubt that in these forms it has an optic function. For this reason the structure wherever found has been regarded as a rudimentary eye, and the "pineal eye" has been called the "unpaired median eye of chordate" animals.
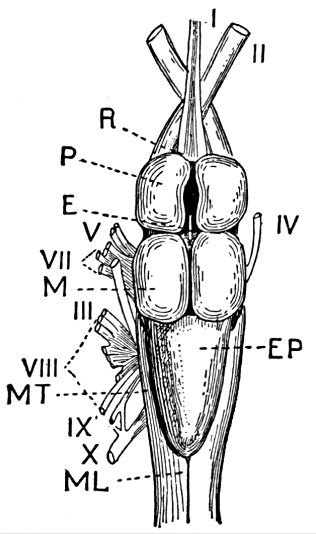
Fig. 81.—Brain of a Perch, Perca flavescens. (After Dean.)
- R. Olfactory lobe.
- P. Cerebrum (prosencephalon).
- E. Epiphysis.
- M. Optic lobes (mesencephalon).
- EP. Cerebellum (epencephalon).
- ML. Medulla oblongata (metencephalon).
- I. First cranial nerve.
- II. Second cranial nerve.
- IV. Fourth cranial nerve.
- V. Fifth cranial nerve.
- VII. Seventh cranial nerve.
- VIII. Eighth cranial nerve.
- IX. Ninth cranial nerve.
- X. Tenth cranial nerve.
Fig. 82.—Petromyzon marinus unicolor (Dekay). Head of Lake Lamprey, showing pineal body. (After Gage.)
It has been supposed that this eye, once possessed by all vertebrate forms, has been gradually lost with the better development of the paired eyes, being best preserved in reptiles as "an outcome of the life-habit which concealed the animal in sand or mud, and allowed the forehead surface alone to protrude, the median eye thus preserving its ancestral value in enabling the animal to look directly upward and backward." This theory receives no support from the structures seen in the fishes.
In none of the fishes is the epiphysis more than a nervous enlargement, and neither in fishes nor in amphibia is there the slightest suggestion of its connection with vision. It seems probable, as suggested by Hertwig and maintained by Dean that the original function of the pineal body was a nervous one and that its connection with or development into a median eye in lizards was a modification of a secondary character. On consideration of the evidence, Dr. Dean concludes that "the pineal structures of the true fishes do not tend to confirm the theory that the epiphysis of the ancestral vertebrates was connected with a median unpaired eye. It would appear, on the other hand, that both in their recent and fossil forms the epiphysis was connected in its median opening with the innervation of the sensory canals of the head. This view seems essentially confirmed by ontogeny. The fact that three successive pairs of epiphyseal outgrowths have been noted in the roof of the thalamencephalon[6] appears distinctly adverse to the theory of a median eye."[7]
The Brain of Primitive Fishes.—The brain of the hagfish differs widely from that of the higher fishes, and the homologies of the different parts are still uncertain. The different ganglia are all solid and are placed in pairs. It is thought that the cerebellum is wanting in these fishes, or represented by a narrow commissure (corpus restiforme) across the front of the medulla. In the lamprey the brain is more like that of the ordinary fish.
In the lancelet there is no trace of brain, the band-like spinal cord tapering toward either end.
The Spinal Cord.—The spinal cord extends from the brain to the tail, passing through the neural arches of the different vertebræ when these are developed. In the higher fishes it is cylindrical and inelastic. In a few fishes (headfish, trunkfish) in which the posterior part of the body is shortened or degenerate, the spinal cord is much shortened, and replaced behind by a structure called cauda equina. In the headfish it has shrunk into "a short and conical appendage to the brain." In the Cyclostomes and chimæra the spinal cord is elastic and more or less flattened or band-like, at least posteriorly.
The Nerves.—The nerves of the fish correspond in general in place and function with those of the higher animals. They are, however, fewer in number, both large nerve-trunks and smaller nerves being less developed than in higher forms.
The olfactory nerves, or first pair, extend through the ethmoid bone to the nasal cavity, which is typically a blind sac with two roundish openings, but is subject to many variations. The optic nerves, or second pair, extend from the eye to the base of the optic lobes. In Cyclostomes these nerves run from each eye to the lobe of its own side. In the bony fishes, or Teleostei, each runs from the eye to the lobe of the opposite side. In the sharks, rays, chimæras, and Ganoids the two optic nerves are joined in a chiasma as in the higher vertebrates.
Other nerves arising in the brain are the third pair, or nervus oculorum motorius, and the fourth pair, nervus trochlearis, both of which supply the muscles of the eye. The fifth pair, nervus trigeminus, and the seventh pair, nervus facialis, arise from the medulla oblongata and are very close together. Their various branches, sensory and motor, ramify among the muscles and sensory areas of the head. The sixth pair, nervus abducens, passes also to muscles of the eye, and in sharks to the nictitating membrane or third eyelid.
The eighth pair, nervus acousticus, leads to the ear. The ninth pair, glosso-pharyngeal, passes to the tongue and pharynx, and forms a ganglion connected with the sympathetic system. The tenth pair, nervus vagus, or pneumogastric nerve, arises from strong roots in the corpus restiforme and the lower part of the medulla oblongata. Its nerves, motor and sensory, reach the muscles of the gill-cavity, heart, stomach, and air-bladder, as well as the muscular system and the skin. In fishes covered with bony plates the skin may be nearly or quite without sensory nerves. The eleventh pair, nervus accessorius, and twelfth pair, nervus hypoglossus, are wanting in fishes.
The spinal nerves are subject to some special modifications, but in the main correspond to similar structures in higher vertebrates. The anterior root of each nerve is without ganglionic enlargement and contains only motor elements. The posterior or dorsal root is sensory only and widens into a ganglionic swelling near the base.
A sympathetic system corresponding to that in the higher vertebrates is found in all the Teleostei, or bony fishes, and in the body of sharks and rays in which it is not extended to the head.
FOOTNOTES:
[6] The thalamencephalon or the interbrain is a name given to the region of the optic thalami, between the bases of the optic lobes and cerebrum.
[7] Fishes Recent and Fossil, p. 55.
CHAPTER VIII
THE ORGANS OF SENSE

The Organs of Smell.—The sense-organs of the fish correspond in general to those of the higher vertebrates. The sense of taste is, however, feeble or wanting, and that of hearing is muffled and without power of acute discrimination, if indeed it exists at all. According to Dr. Kingsley (Vert. Zool., p. 75), "recent experiments tend to show that in fishes the ears are without auditory functions and are solely organs of equilibration."
The sense of smell resides in the nostrils, which have no relation to the work of breathing. No fish breathes through its nostrils, and only in a few of the lowest forms (hagfishes) does the nostril pierce through the roof of the mouth. In the bony fishes the nostril is a single cavity, on either side, lined with delicate or fringed membrane, well provided with blood-vessels, and with nerves from the olfactory lobe. In most cases each nasal cavity has two external openings. These may be simple, or the rim of the nostril may be elevated, forming a papilla or even a long barbel. Either nostril may have a papilla or barbel, or the two may unite in one structure with two openings or with sieve-like openings, or in some degenerate types (Tropidichthys) with no obvious openings at all, the olfactory nerves spreading over the skin of a small papilla. The openings may be round, slit-like, pore-like, or may have various other forms. In certain families of bony fishes (Pomacentridæ, Cichlidæ, Hexagrammidæ), there is but one opening to each nostril. In the sharks, rays, and chimæras there is also but one opening on either side and the nostril is large and highly specialized, with valvular flaps controlled by muscles which are said to enable them "to scent actively as well as to smell passively."
In the lancelet there is a single median organ supposed to be a nostril, a small depression at the front of the head, covered by ciliated membrane. In the hagfish the single median nostril pierces the roof of the mouth, and is strengthened by cartilaginous rings, like those of the windpipe. In the lamprey the single median nostril leads to a blind sac. In the Barramunda (Neoceratodus) there are both external and internal nares, the former being situated just within the upper lip. In all other fishes there is a nasal sac on either side of the head. This has usually, but not always, two openings.
There is little doubt that the sense of smell in fishes is relatively acute, and that the odor of their prey attracts them to it. It is known that flesh, blood, or a decaying carcass will attract sharks, and other predatory fish are drawn in a similar manner. At the same time the strength of this function is yet to be tested by experiments.
Fig. 83.—Dismal Swamp Fish, Chologaster cornutus Agassiz. Supposed ancestor of Typhlichthys. Virginia.
Fig. 84.—Blind Cavefish, Typhlichthys subterraneus Girard. Mammoth Cave, Kentucky.
The Organs of Sight.—The eyes of fishes differ from those of the higher vertebrates mainly in the spherical form of the crystalline lens. This extreme convexity is necessary because the lens itself is not very much denser than the fluid in which the fishes live. The eyes vary very much in size and somewhat in form and position. They are larger in fishes living at a moderate depth than in shore fishes or river fishes. At great depths, as a mile or more, where all light is lost, they may become aborted or rudimentary, and may be covered by the skin. Often species with very large eyes, making the most of a little light or of light from their own luminous spots, will inhabit the same depths with fishes having very small eyes or eyes apparently useless for seeing, retained as vestigial structures through heredity. Fishes which live in caves become also blind, the structures showing every possible phase of degradation. The details of this gradual loss of eyes, whether through reversed selection or hypothetically through inheritance of atrophy produced by disuse, have been given in a number of memoirs on the blind fishes of the Mississippi Valley by Dr. Carl H. Eigenmann.
In some fishes the eye is raised on a short, fleshy stalk and can be moved about at the will of the fish. It is said that the vision of the pond-skipper, Periophthalmus, when hunting insects on the mud flats of Japan or India is "quite equal to that of a frog." It is known also that trout possess keen eyesight, and that they show a marked preference for one sort or another of real or artificial fly. Nevertheless the vision of fishes in general is probably not very precise. They apparently notice motion rather than outline, changes rather than objects, while the extreme curvature of the crystalline lens would seem to render them all near-sighted.

Fig. 85.—Four-eyed Fish, Anableps dovii Gill. Tehuantepec, Mexico.
In the eyes of the fishes there is no lachrymal gland. True eyelids no fishes possess; the integuments of the head pass over the eye, becoming transparent as they cross the orbit. In some fishes part of this integument is thickened, covering the eye fully although still transparent. This forms the adipose eyelid characteristic of the mullet, mackerel, and ladyfish. Many of the sharks possess a distinct nictitating membrane or special eyelid, moved by a set of muscles. The iris in most fishes surrounds a round pupil without much power of contraction. It is frequently brightly colored, red, orange, black, blue, or green. In fishes, like rays or flounders, which lie on the bottom, a dark lobe covers the upper part of the pupil—a curtain to shut out light from above. The cornea is little convex, leaving small space for aqueous humor. In two genera of fishes, Anableps, Dialommus, the cornea is divided by a horizontal partition into two parts. This arrangement permits these fishes, which swim at the surface of the water, to see both in and out of the medium. Anableps, the four-eyed fish, is a fresh-water fish of tropical America, which swims at the surface like a top-minnow, feeding on insects. Dialommus is a marine blenny from the Panama region, apparently of similar habit.
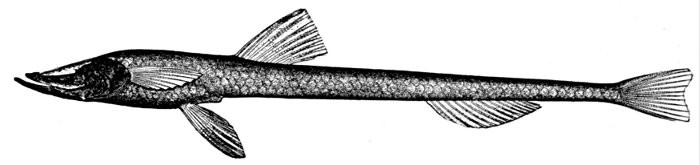
Fig. 86.—Ipnops murrayi Günther.
In one genus of deep-sea fishes, Ipnops, the eyes are spread out to cover the whole upper surface of the head, being modified as luminous areas. Whether these fishes can see at all is not known.

Fig. 87.—Pond-skipper, Boleophthalmus chinensis (Osbeck). Bay of Tokyo, Japan; from nature. K. Morita. (Eye-stalks shrunken in preservation.)
The position of the optic nerves is described in a previous chapter.
In ordinary fishes there is one eye on each side of the head, but in the flounders, by a distortion of the cranium, both appear on the same side. This side is turned uppermost as the fish swims in the water or when it lies on the bottom. This distortion is a matter of development. The very young flounder swims with its broad axis vertical in the water, and it has one eye on either side. As soon as it rests on the bottom it begins to lean to one side. The lower eye changes its axis and by degrees travels across the face of the fish, part of the bony interorbital moving with it across to the other side. In some soles it is said to pass through the substance of the head, reappearing on the other side. In all species which the writer has examined the cranium is twisted, the eye moving with the bones; and the frontal bone is divided, a new orbit being formed by this division. In most northern flounders the eyes are on the right side in the adult, in tropical forms more frequently on the left, these distinctions corresponding with others in the structure of the fish.
In the lowest of the fish-like forms, the lancelet, the eye is simply a minute pigment-spot situated in the anterior wall of the ventricle at the anterior end of the central nervous system. In the hagfishes, which stand next highest in the series, the eye, still incomplete, is very small and hidden by the skin and muscles. This condition is very different from that of the blind fishes of the higher groups, in which the eye is lost through atrophy, because in life in caves or under rocks the function of seeing is no longer necessary.
The Organs of Hearing.—The ear of the typical fish consists of the labyrinth only, including the vestibule and usually three semicircular canals, these dilating into sacs which contain one or more large, loose bones, the ear-stones or otoliths. In the lampreys there are two semicircular canals, in the hagfish but one. There is no external ear, no tympanum, and no Eustachian tube. The ear-sac on each side is lodged in the skull or at the base of the cranial cavity. It is externally surrounded by bone or cartilage, but sometimes it lies near a fontanelle or opening in the skull above. In some fishes it is brought into very close connection with the anterior end of the air-bladder. The latter organ it is thought may form part of the apparatus for hearing. The arrangement for this purpose is especially elaborate in the carp and the catfish families. In these fishes and their relatives (called Ostariophysi) the two vestibules are joined in a median sac (sinus impar) in the substance of the basioccipital. This communicates with two cavities in the atlas, which again are supported by two small bones, these resting on a larger one in connection with the front of the air-bladder. The system of bones is analogous to that found in the higher vertebrates, but it connects with the air-bladder, not with an external tympanum. The bones are not homologous with those of the ear of higher animals, being processes of the anterior vertebræ. The tympanic chain of higher vertebrates has been thought homologous with the suspensory of the mandible.

Fig. 88.—Brook Lamprey, Lampetra wilderi Jordan and Evermann. (After Gage.) Cayuga Lake.
The otoliths, commonly two in each labyrinth, are usually large, firm, calcareous bodies, with enamelled surface and peculiar grooves and markings. Each species has its own form of otolith, but they vary much in different groups of fishes.
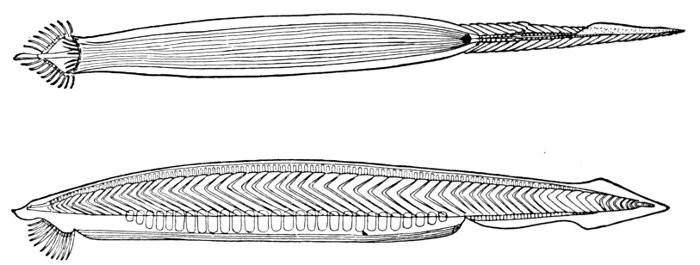
Fig. 89.—European Lancelet, Branchiostoma lanceolatum (Pallas). (After Parker and Haswell.)
In the Elasmobranchs (sharks and rays) and in the Dipnoans the ear-sac is enclosed in the cartilaginous substance of the skull. There is a small canal extending to the surface of the skull, ending sometimes in a minute foramen. The otoliths in these fishes are soft and chalk-like.
The lancelet shows no trace of an ear. In the cyclostomes, hagfishes, and lampreys it forms a capsule of relatively simple structure conspicuous in the prepared skeleton.
The sense of hearing in fishes cannot be very acute, and is at the most confined to the perception of disturbances in the water. Most movements of the fish are governed by sight rather than by sound. It is in fact extremely doubtful whether fishes really hear at all, in a way comparable to the auditory sense in higher vertebrates. Recent experiments of Professor G. H. Parker on the killifish tend to show a moderate degree of auditory sense which grades into the sense of touch, the tubes of the lateral line assisting in both hearing and touch. While the killifish responds to a bass-viol string, there may be some fishes wholly deaf.
Voices of Fishes.—Some fishes make distinct noises variously described as quivering, grunting, grating, or singing. The name grunt is applied to species of Hæmulon and related genera, and fairly describes the sound these fishes make. The Spanish name ronco or roncador (grunter or snorer) is applied to several fishes, both sciænoid and hæmuloid. The noise made by these fishes may be produced by forcing air from part to part of the complex air-bladder, or it may be due to grating one on another of the large pharyngeals. The grating sounds arise, no doubt, from the pharyngeals, while the quivering or singing sounds arise in the air-bladder. The midshipman, Porichthys notatus, is often called singing fish, from a peculiar sound it emits. These sounds have not yet been carefully investigated.
The Sense of Taste.—It is not certain that fishes possess a sense of taste, and it is attributed to them only through their homology with the higher animals. The tongue is without delicate membranes or power of motion. In some fishes certain parts of the palate or pharyngeal region are well supplied with nerves, but no direct evidence exists that these have a function of discrimination among foods. Fishes swallow their food very rapidly, often whole, and mastication, when it takes place, is a crushing or cutting process, not one likely to be affected by the taste of the food.
The Sense of Touch.—The sense of touch is better developed among fishes. Most of them flee from contact with actively moving objects. Many fishes use sensitive structures as a means of exploring the bottom or of feeling their way to their food. The barbel or fleshy filament wherever developed is an organ of touch. In some fishes, barbels are outgrowths from the nostrils. In the catfish the principal barbel grows from the rudimentary maxillary bone. In the horned dace and gudgeon the little barbel is attached to the maxillary. In other fishes barbels grow from the skin of the chin or snout. In the goatfish and surmullet the two chin barbels are highly specialized. In Polymixia the chin barbels are modified branchiostegals. In the codfish the single beard is little developed. In the gurnards and related forms the lower rays of the pectoral are separate and barbel-like. Detached rays of this sort are found in the thread-fins (Polynemidæ), the gurnards (Triglidæ), and in various other fishes. Barbels or fleshy flaps are often developed over the eyes and sometimes on the scales or the fins.

Fig. 90.—Goatfish, Pseudupeneus maculatus (Bloch). Woods Hole.
The structure of the lateral line and its probable relation as a sense-organ is discussed on page 23. It is probable that it is associated with sense of touch, and hearing as well, the internal ear being originally "a modified part of the lateral-line system," as shown by Parker,[8] who calls the skin the lateral line and the ear "three generations of sense-organs."
The sense of pain is very feeble among fishes. A trout has been known to bite at its own eye placed on a hook, and similar insensibility has been noted in the pike and other fishes. "The Greenland shark, when feeding on the carcass of a whale, allows itself to be repeatedly stabbed in the head without abandoning its prey." (Günther.)
FOOTNOTES:
[8] See Parker, on the sense of hearing in fishes, American Naturalist for March, 1903.
CHAPTER IX
THE ORGANS OF REPRODUCTION

The Germ-cells.—In most fishes the germ-cells are produced in large sacs, ovaries or testes, arranged symmetrically one on either side of the posterior part of the abdominal cavity. The sexes are generally but not always similar externally, and may be distinguished on dissection by the difference between the sperm-cells and the ova. The ovary with its eggs is more yellow in color and the contained cells appear granular. The testes are whitish or pinkish, their secretion milk-like, and to the naked eye not granular.
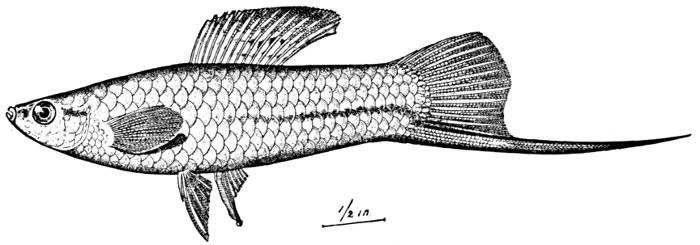
Fig. 91.—Sword-tail Minnow, male, Xiphophorus helleri Heckel. The anal fin modified as an intromittent organ. Vera Cruz.
In a very few cases both organs have been found in the same fish, as in Serranus, which is sometimes truly hermaphrodite. All fishes, however, seem to be normally diœcious, the two sexes in different individuals. Usually there are no external genital organs, but in some species a papilla or tube is developed at the end of the urogenital sinus. This may exist in the breeding season only, as in the fresh-water lampreys, or it may persist through life as in some gobies. In the Elasmobranchs, cartilaginous claspers, attached to the ventral fins in the male, serve as a conduit for the sperm-cells.
The Eggs of Fishes.—The great majority of fishes are oviparous, the eggs being fertilized after deposition. The eggs are laid in gravel or sand or other places suitable for the species, and the milt containing the sperm-cells of the male is discharged over or among them in the water. A very small quantity of the sperm-fluid may impregnate a large number of eggs. But one sperm-cell can enter a particular egg. In a number of families the species are ovoviviparous, the eggs being hatched in the ovary or in a dilated part of the oviduct, the latter resembling a real uterus. In some sharks there is a structure analogous to the placenta of higher animals, but not of the same structure or origin. In the case of viviparous fishes actual copulation takes place and there is usually a modification of some organ to effect transfer of the sperm-cells. This is the purpose of the sword-shaped anal fin in many top-minnows (Pæciliidæ), the fin itself being placed in advance of its usual position. In the surf-fishes (Embiotocidæ) the structure of part of the anal fin is modified, although it is not used as an intromittent organ. In the Elasmobranchs, as already stated, large organs of cartilage (claspers) are developed from the ventral fins.
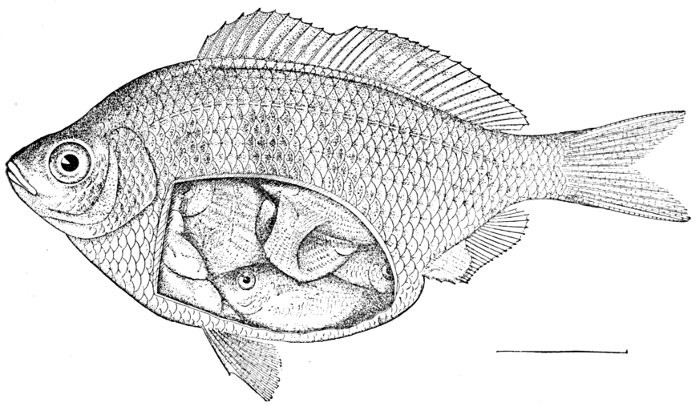
Fig. 92.—White Surf-fish, viviparous, with young, Cymatogaster aggregatus Gibbons. San Francisco.
In some viviparous fishes, as in the rockfishes (Sebastodes) and rosefishes (Sebastes), the young are very minute at birth.

Fig. 93.—Goodea luitpoldi (Steindachner). A viviparous fish from Lake Patzcuaro, Mexico. Family Pæciliidæ. (After Meek.)
In others, as the surf-fishes (Embiotocidæ), they are relatively large and few in number. In the viviparous sharks, which constitute the majority of the species of living sharks, the young are large at birth and prepared to take care of themselves.

Fig. 94.—Egg of Callorhynchus antarcticus, the Bottle-nosed Chimæra. (After Parker and Haswell.)
The eggs of fishes vary very much in size and form. In those sharks and rays which lay eggs the ova are deposited in a horny egg-case, in color and texture suggesting the kelp in which they are laid. The eggs of the bullhead sharks (Heterodontus) are spirally twisted, those of the cat-sharks (Scyliorhinidæ) are quadrate with long filaments at the angles. Those of rays are wheelbarrow-shaped with four "handles." One egg-case of a ray may sometimes contain several eggs and develop several young. The eggs of lancelets are small, but those of the hagfishes are large, ovate, with fibres at each side, each with a triple hook at tip. The chimæra has also large egg-cases, oblong in form.

Fig. 95.—Egg of the Hagfish, Myxine limosa Girard, showing threads for attachment. (After Dean.)
In the higher fishes the eggs are spherical, large or small according to the species, and varying in the firmness of their outer walls. All contain food-yolk from which the embryo in its earlier stages is fed. The eggs of the eel (Anguilla) are microscopic. According to Günther 25,000 eggs have been counted in the herring, 155,000 in the lumpfish, 3,500,000 in the halibut, 635,200 in the sturgeon, and 9,344,000 in the cod. Smaller numbers are found in fishes with large ova. The red salmon has about 3500 eggs, the king salmon about 5200. Where an oviduct is present the eggs are often poured out in glutinous masses, as in the bass. When, as in the salmon, there is no oviduct, the eggs lie separate and do not cohere together. It is only with the latter class of fishes, those in which the eggs remain distinct, that artificial impregnation and hatching is practicable. In this regard the value of the salmon and trout is predominant. In some fishes, especially those of elongate form, as the needle-fish (Tylosurus), the ovary of but one side is developed.
Fig. 96.—Egg of Port Jackson Shark, Heterodontus philippi (Lacépède). (After Parker and Haswell.)
Protection of the Young.—In most fishes the parents take no care of their eggs or young. In some catfishes (Platystacus) the eggs adhere to the under surface of the female. In a kind of pipefish (Solenostomus), a large pouch for retention of the eggs is formed on the belly of the female. In the sea-horses and pipefishes a pouch is formed in the skin, usually underneath the tail of the male. Into this the eggs are thrust, and here the young fishes hatch out, remaining until large enough to take care of themselves. In certain sea catfishes (Galeichthys, Conorhynchos) the male carries the eggs in his mouth, thus protecting them from the attacks of other fishes. In numerous cases the male constructs a rough nest, which he defends against all intruders, against the female as well as against outside enemies. The nest-building habit is especially developed in the sticklebacks (Gasterosteidæ), a group in which the male fish, though a pygmy in size, is very fierce in disposition.
In a minnow of Europe (Rhodeus amarus) the female is said to deposit her eggs within the shells of river mussels.
Sexual Modification.—In the relatively few cases in which the sexes are unlike the male is usually the brighter in color and with more highly developed fins. Blue, red, black, and silvery-white pigment are especially characteristic of the male, the olivaceous and mottled coloration of the female. Sometimes the male has a larger mouth, or better developed crests, barbels, or other appendages. In some species the pattern of coloration in the two sexes is essentially different.
In various species the male develops peculiar structures not found in the female, and often without any visible purpose. In the chimæra a peculiar cartilaginous hook armed with a brush of enamelled teeth at the tip is developed on the forehead in the male only. In the skates or true rays (Raja) the pectoral fin has near its edge two rows of stout incurved spines. These the female lacks. In the breeding season, among certain fishes, the male sometimes becomes much brighter by the accumulation of bright red or blue pigment accompanied by black or white pigment cells. This is especially true in the minnows (Notropis), the darters (Etheostoma), and other fresh-water species which spawn in the brooks of northern regions in the spring. In the minnows and suckers horny excrescences are also developed on head, body, or fins, to be lost after the deposition of the spawn.
In the salmon, especially those of the Pacific, the adult male becomes greatly distorted in the spawning season, the jaws and teeth being greatly elongated and hooked or twisted so that the fish cannot shut its mouth. The Atlantic salmon and the trout show also some elongation of the jaws, but not to the same extent.
In those fishes which pair the relation seems not to be permanent, nor is there anything to be called personal affection among them so far as the writer has noticed.
There is no evidence that the bright colors or nuptial adornments of the males are enhanced by sexual selection. In most species the males deposit the sperm-cells in spawning-grounds without much reference to the preference of the females. In general the brightest colors are not found among viviparous fishes. None of the groups in which the males are showily colored, while the females are plain, belong to this class. The brightest colors are found on the individuals most mature or having greatest vitality.
CHAPTER X
EMBRYOLOGY AND GROWTH OF FISHES

Segmentation of the Egg.—The egg of the fish develops only after fertilization (amphimixis). This process is the union of its nuclear substance with that of the sperm-cell from the male, each cell carrying its equal share in the function of heredity. When this process takes place the egg is ready to begin its segmentation. The eggs of all fishes are single cells containing more or less food-yolk. The presence of this food-yolk affects the manner of segmentation in general, those eggs having the least amount of food-yolk developing most typically. The simplest of all fish like vertebrates, the lancelet (Branchiostoma) has very small eggs, and in their early development it passes through stages that are typical for all many-celled animals. The first stage in development is the simple splitting of the egg into two halves. These two daughter cells next divide so that there are four cells; each of these divides, and this division is repeated until a great number of cells is produced. The phenomenon of repeated division of the germ-cell is called cleavage, and this cleavage is the first stage of development in the case of all many-celled animals. Instead of forming a solid mass the cells arrange themselves in such a way as to form a hollow ball, the wall being a layer one cell thick. The included cavity is called the segmentation cavity, and the whole structure is known as a blastula. This stage also is common to all the many-celled animals. The next stage is the conversion of the blastula into a double-walled cup, known as a gastrula by the pushing in of one side. All the cells of the blastula are very small, but those on one side are somewhat larger than those of the other, and here the wall first flattens and then bends in until finally the larger cells come into contact with the smaller and the segmentation cavity is entirely obliterated. There is now an inner layer of cells and an outer layer, the inner layer being known as the endoblast and the outer as the ectoblast. The cavity of the cup thus formed is the archenteron and gives rise primarily to the alimentary canal. This third well-marked stage is called the gastrula stage; and it is thought to occur either typically or in some modified form in the development of all metazoa, or many-celled animals. In the lampreys, the Ganoids, and the Dipnoans the eggs contain a much greater quantity of yolk than those of the lancelet, but the segmentation resembles that of the lancelet in that it is complete; that is, the whole mass of the egg divides into cells. There is a great difference, however, in the size of the cells, those at the upper pole being much smaller than those at the lower. In Petromyzon and the Dipnoans blastula and gastrula stages result, which, though differing in some particulars from the corresponding stages of the lancelet, may yet readily be compared with them. In the hagfishes, sharks, rays, chimæras, and most bony fishes there is a large quantity of yolk, and the protoplasm, instead of being distributed evenly throughout the egg, is for the most part accumulated upon one side, the nucleus being within this mass of protoplasm. When the food substance or yolk is consumed and the little fish is able to shift for itself, it leaves the egg-envelopes and is said to be hatched. The figures on page 135 show some of the stages by which cells are multiplied and ultimately grouped together to form the little fish.
Post-embryonic Development.—In all the fishes the development of the embryo goes on within the egg long after the gastrula stage is passed, and until the embryo becomes a complex body, composed of many differing tissues and organs. Almost all the development may take place within the egg, so that when the young animal hatches there is necessary little more than a rapid growth and increase of size to make it a fully developed mature animal. This is the case with most fishes: a little fish just hatched has most of the tissues and organs of a full-grown fish, and is simply a small fish. But in the case of some fishes the young hatches from the egg before it has reached such an advanced state of development, and the young looks very different from its parent. It must yet undergo considerable change before it reaches the structural condition of a fully developed and fully grown fish. Thus the development of most fishes is almost wholly embryonic development—that is, development within the egg or in the body of the mother—while the development of some of them is to a considerable degree post-embryonic or larval development. There is no important difference between embryonic and post-embryonic development. The development is continuous from egg-cell to mature animal and, whether inside or outside of an egg, it goes on with a degree of regularity. While certain fishes are subject to a sort of metamorphosis, the nature of this change is in no way to be compared with the change in insects which undergo a complete metamorphosis. In the insects all the organs of the body are broken down and rebuilt in the process of change. In all fishes a structure once formed maintains a more nearly continuous integrity although often considerably altered in form.
General Laws of Development.—The general law of development may be briefly stated as follows: All many-celled animals begin life as a single cell, the fertilized egg-cell; each animal goes through a certain orderly series of developmental changes which, accompanied by growth, leads the animal to change from single-cell to many-celled, complex form characteristic of the species to which the animal belongs; this development is from simple to complex structural condition; the development is the same for all individuals of one species. While all animals begin development similarly, the course of development in the different groups soon diverges, the divergence being of the nature of a branching, like that shown in the growth of a tree. In the free tips of the smallest branches we have represented the various species of animals in their fully developed condition, all standing clearly apart from each other. But in tracing back the development of any kind of animal we soon come to a point where it very much resembles or becomes apparently identical with some other kind of animal, and going farther back we find it resembling other animals in their young condition, and so on until we come to that first stage of development, that trunk stage where all animals are structurally alike. Any animal at any stage in its existence differs absolutely from any other kind of animal, in this respect: it can develop into only its own kind. There is something inherent in each developing animal that gives it an identity of its own. Although in its young stages it may be indistinguishable from some other species of animal in its young stages, it is sure to come out, when fully developed, an individual of the same kind as its parents were or are. The young fish and the young salamander may be alike to all appearance, but one embryo is sure to develop into a fish, and the other into a salamander. This certainty of an embryo to become an individual of a certain kind is called the law of heredity. Viewed in the light of development, there must be as great a difference between one egg and another as between one animal and another, for the greater difference is included in the less.
The Significance of Facts of Development.—The significance of the process of development in any species is yet far from completely understood. It is believed that many of the various stages in the development of an animal correspond to or repeat the structural condition of the animal's ancestors. Naturalists believe that all animals having a notochord at any stage in their existence are related to each other through being descended from a common ancestor, the first or oldest chordate or back-boned animal. In fact it is because all these chordate animals—the lancelets, lampreys, fishes, batrachians, the reptiles, the birds, and the mammals—have descended from a common ancestor that they all develop a notochord, and those most highly organized replace this by a complete back-bone. It is believed that the descendants of the first back-boned animal have, in the course of many generations, branched off little by little from the original type until there came to exist very real and obvious differences among the back-boned animals—differences which among the living back-boned animals are familiar to all of us. The course of development of an individual animal is believed to be a very rapid and evidently much condensed and changed recapitulation of the history which the species or kind of animal to which the developing individual belongs has passed through in the course of its descent through a long series of gradually changing ancestors. If this is true, then we can readily understand why the fish and the salamander and the tortoise and bird and rabbit are all alike in their earlier stages of development, and gradually come to differ more and more as they pass through later and later developmental stages.
Development of the Bony Fishes.[9] The mode of development of bony fishes differs in many and apparently important regards from that of their nearest kindred, the Ganoids. In their eggs a large amount of yolk is present, and its relations to the embryo have become widely specialized. As a rule, the egg of a Teleost is small, perfectly spherical, and enclosed in delicate but greatly distended membranes. The germ disc is especially small, appearing on the surface as an almost transparent fleck. Among the fishes whose eggs float at the surface during development, as of many pelagic Teleosts, e.g., the sea-bass, Centropristes striatus, the yolk is lighter in specific gravity than the germ; it is of fluid-like consistency, almost transparent. In the yolk at the upper pole of the egg an oil globule usually occurs; this serves to lighten the relative weight of the entire egg, and from its position must aid in keeping this pole of the egg uppermost.
The Origin of the Lungs.—The following account of the function of the air-bladder and of its development and decline is condensed from an article by Mr. Charles Morris:[5]
[6] The thalamencephalon or the interbrain is a name given to the region of the optic thalami, between the bases of the optic lobes and cerebrum.
[7] Fishes Recent and Fossil, p. 55.
In none of the fishes is the epiphysis more than a nervous enlargement, and neither in fishes nor in amphibia is there the slightest suggestion of its connection with vision. It seems probable, as suggested by Hertwig and maintained by Dean that the original function of the pineal body was a nervous one and that its connection with or development into a median eye in lizards was a modification of a secondary character. On consideration of the evidence, Dr. Dean concludes that "the pineal structures of the true fishes do not tend to confirm the theory that the epiphysis of the ancestral vertebrates was connected with a median unpaired eye. It would appear, on the other hand, that both in their recent and fossil forms the epiphysis was connected in its median opening with the innervation of the sensory canals of the head. This view seems essentially confirmed by ontogeny. The fact that three successive pairs of epiphyseal outgrowths have been noted in the roof of the thalamencephalon[6] appears distinctly adverse to the theory of a median eye."[7]
In none of the fishes is the epiphysis more than a nervous enlargement, and neither in fishes nor in amphibia is there the slightest suggestion of its connection with vision. It seems probable, as suggested by Hertwig and maintained by Dean that the original function of the pineal body was a nervous one and that its connection with or development into a median eye in lizards was a modification of a secondary character. On consideration of the evidence, Dr. Dean concludes that "the pineal structures of the true fishes do not tend to confirm the theory that the epiphysis of the ancestral vertebrates was connected with a median unpaired eye. It would appear, on the other hand, that both in their recent and fossil forms the epiphysis was connected in its median opening with the innervation of the sensory canals of the head. This view seems essentially confirmed by ontogeny. The fact that three successive pairs of epiphyseal outgrowths have been noted in the roof of the thalamencephalon[6] appears distinctly adverse to the theory of a median eye."[7]
The Lateral Line as a Mucous Channel.—The more primitive condition of the lateral line is seen in the sharks and chimæras, in which fishes it appears as a series of channels in or under the skin. These channels are filled with mucus, which exudes through occasional open pores. In many fishes the bones of the skull are cavernous, that is, provided with cavities filled with mucus. Analogous to these cavities are the mucous channels which in primitive fishes constitute the lateral line.
[8] See Parker, on the sense of hearing in fishes, American Naturalist for March, 1903.
The structure of the lateral line and its probable relation as a sense-organ is discussed on page 23. It is probable that it is associated with sense of touch, and hearing as well, the internal ear being originally "a modified part of the lateral-line system," as shown by Parker,[8] who calls the skin the lateral line and the ear "three generations of sense-organs."
The Significance of Facts of Development.—The significance of the process of development in any species is yet far from completely understood. It is believed that many of the various stages in the development of an animal correspond to or repeat the structural condition of the animal's ancestors. Naturalists believe that all animals having a notochord at any stage in their existence are related to each other through being descended from a common ancestor, the first or oldest chordate or back-boned animal. In fact it is because all these chordate animals—the lancelets, lampreys, fishes, batrachians, the reptiles, the birds, and the mammals—have descended from a common ancestor that they all develop a notochord, and those most highly organized replace this by a complete back-bone. It is believed that the descendants of the first back-boned animal have, in the course of many generations, branched off little by little from the original type until there came to exist very real and obvious differences among the back-boned animals—differences which among the living back-boned animals are familiar to all of us. The course of development of an individual animal is believed to be a very rapid and evidently much condensed and changed recapitulation of the history which the species or kind of animal to which the developing individual belongs has passed through in the course of its descent through a long series of gradually changing ancestors. If this is true, then we can readily understand why the fish and the salamander and the tortoise and bird and rabbit are all alike in their earlier stages of development, and gradually come to differ more and more as they pass through later and later developmental stages.
[9] This account of the normal development of the Teleost fishes is condensed from Dr. Dean's "Fishes Living and Fossil," in which work the details of growth in the Teleost are contrasted with those of other types of fishes.
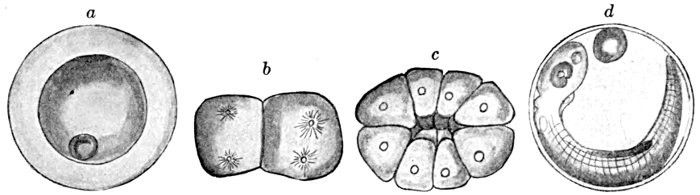
Fig. 97.—Development of Sea-bass, Centropristes striatus (Linnæus). a, egg prior to germination; b, germ-disk after first cleavage; c, germ-disk after third cleavage; d, embryo just before hatching. (After H. V. Wilson.)
In the early segmentation of the germ the first cleavage plane is established, and the nuclear divisions have taken place for the second; in the latter the third cleavage has been completed. As in other fishes these cleavages are vertical, the third parallel to the first. A segmentation cavity occurs as a central space between the blastomeres, as it does in the sturgeon and garpike.
In stages of late segmentation the segmentation cavity is greatly flattened, but extends to the marginal cells of the germ-disk; its roof consists of two tiers of blastomeres, its floor of a thin film of the unsegmented substance of the germ; the marginal blastomeres are continuous with both roof and floor of the cavity, and are produced into a thin film which passes downward, around the sides of the yolk. Later the segmentation cavity is still further flattened; its roof is now a dome-shaped mass of blastomeres; the marginal cells have multiplied, and their nuclei are seen in the layer of the germ, below the plane of the segmentation cavity. These are seen in the surface view of the marginal cells of this stage; they are separated by cell boundaries only at the sides; below they are continuous in the superficial down-reaching layer of the germ. The marginal cells shortly lose all traces of having been separate; their nuclei, by continued division, spread into the layer of germ flooring the segmentation cavity, and into the delicate film of germ which now surrounds the entire yolk. Thus is formed the periblast of the Teleost development, which from this point onward is to separate the embryo from the yolk; it is clearly the specialized inner part of the germ, which, becoming fluid-like, loses its cell-walls, although retaining and multiplying its nuclei. Later the periblast comes into intimate relations with the growing embryo; it lies directly against it, and appears to receive cell increments from it at various regions; on the other hand, the nuclei of the periblast, from their intimate relations with the yolk, are supposed to subserve some function in its assimilation.
Aside from the question of periblast, the growth of the blastoderm appears not unlike that of the sturgeon. From the blastula stage to that of the early gastrula, the changes have been but slight; the blastoderm has greatly flattened out as its margins grow downward, leaving the segmentation cavity apparent. The rim of the blastoderm has become thickened as the 'germ-ring'; and immediately in front of the dorsal lip of the blastopore its thickening marks the appearance of the embryo. The germ-ring continues to grow downward, and shows more prominently the outline of the embryo; this now terminates at the head region; while on either side of this point spreads out tail-ward on either side the indefinite layer of outgrowing
mesoderm. In the next stage the closure of the blastopore
is rapidly becoming completed; in front of it stretches the widened and elongated form of the embryo. The yolk-plug is next replaced by periblast, the dorsal lip by the tail-mass, or more accurately the dorsal section of the germ-rim; the cœlenteron under the dorsal lip has here disappeared, on account of the close approximation of the embryo to the periblast; its last remnant, the Kupffer's vesicle, is shortly to disappear. The germ-layers become confluent, but, unlike the sturgeon, the flattening of the dorsal germ-ring does not permit the formation of a neurenteric canal.

Fig. 98. Sea-bass, Centropristes striatus, natural size. (From life, by R. W. Shufeldt.)—Page 137.
The process of the development of the germ-layers in Teleosts appears as an abbreviated one, although in many of its details it is but imperfectly known. In the development of the medullary groove, as an example, the following peculiarities exist: the medullary region is but an insunken mass of cells without a trace of the groove-like surface indentation. It is only later, when becoming separate from the ectoderm, that it acquires its rounded character; its cellular elements then group themselves symmetrically with reference to a sagittal plane, where later, by their dissociation, the canal of the spinal cord is formed. The growth of the entoderm is another instance of specialized development. In an early stage the entoderm exists in the axial region, its thickness tapering away abruptly on either side; its lower surface is closely apposed to the periblast; its dorsal thickening will shortly become separate as the notochord. In a following stage of development the entoderm is seen to arch upward in the median line as a preliminary stage in the formation of the cavity of the gut. Later, by the approximation of the entoderm-cells in the median ventral line, the condition is reached where the completed gut-cavity exists.
The formation of the mesoderm in Teleosts is not definitely understood. It is usually said to arise as a process of 'delamination,' i.e., detaching itself in a mass from the entoderm. Its origin is, however, looked upon generally as of a specialized and secondary character.
The mode of formation of the gill-slit of the Teleost does not differ from that in other groups; an evagination of the entoderm coming in contact with an invaginated tract of ectoderm fuses, and at this point an opening is later established.
The late embryo of the Teleost, though of rounded form, is the more deeply implanted in the yolk-sac than that of the sturgeon; it is transparent, allowing notochord, primitive segments, heart, and sense-organs to be readily distinguished; at about this stage both anus and mouth are making their appearance.
Fig. 99.—Young Sword-fish, Xiphias gladius (Linnæus). (After Lütken.)
The Larval Development of Fishes.[10]—"When the young fish has freed itself from its egg-membranes it gives but little suggestion of its adult form. It enters upon a larval existence, which continues until maturity. The period of change of form varies widely in the different groups of fishes, from a few weeks' to longer than a year's duration; and the extent of the changes that the larva undergoes are often surprisingly broad, investing every organ and tissue of the body, the immature fish passing through a series of form stages which differ one from the other in a way strongly contrasting with the mode of growth of amniotes; since the chick, reptile, or mammal emerges from its embryonic membranes in nearly its adult form.

Fig. 100.—Sword-fish, Xiphias gladius (Linnæus). (After Day.)
The fish may, in general, be said to begin its existence as a larva as soon as it emerges from its egg-membranes. In some instances, however, it is difficult to decide at what point the larval stage is actually initiated: thus in sharks the excessive amount of yolk material which has been provided for the growth of the larva renders unnecessary the emerging from the egg at an early stage; and the larval period is accordingly to be traced back to stages that are still enclosed in the egg-membranes. In all cases the larval life may be said to begin when the following conditions have been fulfilled: the outward form of the larva must be well defined, separating it from the mass of yolk, its motions must be active, it must possess a continuous vertical fin-fold passing dorsally from the head region to the body terminal, and thence ventrally as far as the yolk region; and the following structures, characteristic in outward appearance, must also be established: the sense-organs—eye, ear, and nose—mouth and anus, and one or more gill-clefts.
Fig. 101.—Larva of the Sail-fish, Istiophorus, very young. (After Lütken.)

Fig. 102.—Larva of Brook Lamprey, Lampetra wilderi, before transformation, being as large as the adult, toothless, and more distinctly segmented.

Fig. 103.—Common Eel. Anguilla chrisypa Rafinesque. Family Anguillidæ.
Among the different groups of fishes the larval changes are brought about in widely different ways. These larval peculiarities appear at first of far-reaching significance, but may ultimately be attributed, the writer believes, to changed environmental conditions, wherein one process may be lengthened, another shortened. So, too, the changes from one stage to another may occur with surprising abruptness. As a rule, it may be said the larval stage is of longest duration in the Cyclostomes, and thence diminished in length in sharks, lung-fishes, Ganoids, and Teleosts; in the last-named group a very much curtailed (i.e., precocious) larval life may often occur.
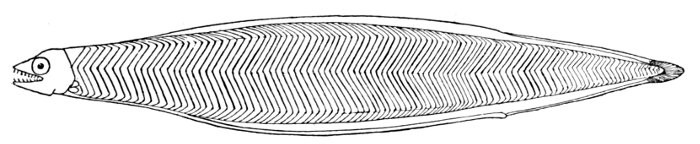
Fig. 104.—Larva of Common Eel, Anguilla chrisypa (Rafinesque), called Leptocephalus grassii. (After Eigenmann.)
The metamorphoses of the newly hatched Teleost must finally be reviewed; they are certainly the most varied and striking of all larval fishes, and, singularly enough, appear to be crowded into the briefest space of time; the young fish, hatched often as early as on the fourth day, is then of the most immature character; it is transparent, delicate, easily injured, inactive; within a month, however, it may have assumed almost every detail of its mature form. A form hatching three millimeters in length may acquire the adult form before it becomes much longer than a centimeter.
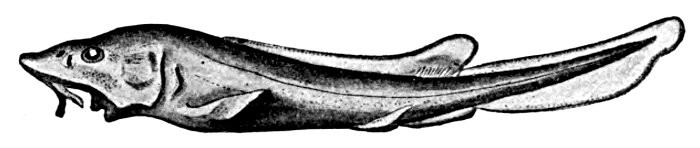
Fig. 105.—Larva of Sturgeon, Acipenser sturio (Linnæus). (After Kupffer, per Dean.)

Fig. 106.—Larva (called Tholichthys) of Chætodon sedentarius (Poey). Cuba. (After Lütken,)

Fig. 107.—Butterfly-fish, Chætodon capistratus Linnæus. Jamaica.
Peculiar Larval Forms.—The young fish usually differs from the adult mainly in size and proportions. The head is larger in the young, the fins are lower, the appendages less developed, and the body more slender in the young than in the adult. But to most of these distinctions there are numerous exceptions, and in some fish there is a change so marked as to be fairly called a metamorphosis. In such cases the young fish in its first condition is properly called a larva. The larva of the lamprey (Petromyzon) is nearly blind and toothless, with slender head, and was long supposed to belong to a different genus (Ammocœtes) from the adult. The larva of sharks and rays, and also of Dipnoans and Crossopterygians, are provided with bushy external gills, which disappear in the process of development. In most soft-rayed fishes the embryonic fringe which precedes the development of the vertical fins persists for a considerable time. In many young fishes, especially the Chætodontidæ and their allies (butterfly-fishes), the young fish has the head armed with broad plates formed by the backward extension of certain membrane-bones. In other forms the bones of the head are in the young provided with long spines or with serrations, which vanish totally with age. Such a change is noticeable in the swordfish. In this species the production of the bones of the snout and upper jaw into a long bony sword, or weapon of offense, takes place only with age. The young fish have jaws more normally formed, and armed with ordinary teeth. In the headfish (Mola mola) large changes take place in the course of growth, and the young have been taken for a different type of fishes. Among certain soft-rayed fishes and eels the young is often developed in a peculiar way, being very soft, translucent, or band-like, and formed of large or loosely aggregated cells. These peculiar organisms, long known as leptocephali, have been shown to be the normal young of fishes when mature very different. In the ladyfish (Albula) Dr. Gilbert has shown, by a full series of specimens, that in their further growth these pellucid fishes shrink in size, acquiring greater compactness of body, until finally reaching about half their maximum length as larvæ. After this, acquiring essentially the form of the adult fish, they begin a process of regular growth. This leptocephalous condition is thought by Günther to be due to arrest of growth in abnormal individuals, but this is not the case in Albula, and it is probably fully normal in the conger and other eels. In the surf-fishes the larvæ have their vertical fins greatly elevated, much higher than in the adult, while the body is much more closely compressed. In the deal-fish (Trachypterus) the form of the body and fins changes greatly with age, the body becoming more elongate and the fins lower. The differences between different stages of the same fish seem greater than the differences between distinct species. In fact with this and with other forms which change with age, almost the only test of species is found in the count of the fin-rays. So far as known the numbers of these structures do not change. In the moonfishes (Carangidæ) the changes with age are often very considerable. We copy Lütken's figure of the changes in the genus Selene (fig. 113). Similar changes take place in Alectis, Vomer, and other genera.

Fig. 108.—Mola mola (Linnæus). Very early larval stage of the Headfish, called Centaurus boöps. (After Richardson.)

Fig. 109.—Mola mola (Linnæus). Early larval stage, called Molacanthus nummularis. (After Ryder.)

Fig. 110.—Mola mola (Linnæus). Advanced larval stage. (After Ryder.)
The Development of Flounders.—In the great group of flounders and soles (Heterosomata) the body is greatly compressed and the species swim on one side or lie flat on the bottom, with one side uppermost. This upper side is colored like the bottom, sand-color, gray, or brown, while the lower side is mostly white. Both eyes are brought around to the upper side by a twisting of the cranium and a modification or division of the frontal bones. When the young flounder is hatched it is translucent and symmetrical, swimming vertically in the water, with one eye on either side of the head. After a little the young fish rests the ventral edge on the bottom. It then leans to one side, and as its position gradually becomes horizontal the eye on the lower side moves across with its frontal and other bones to the other side. In most species it passes directly under the first interneurals of the dorsal fin. These changes are best observed in the genus Platophrys.
Hybridism.—Hybridism is very rare among fishes in a state of nature. Two or three peculiar forms among the snappers (Lutianus) in Cuba seem fairly attributable to hybridism, the single specimen of each showing a remarkable mixture of characters belonging to two other common species. Hybrids may be readily made in artificial impregnation among those fishes with which this process is practicable. Hybrids of the different salmon or trout usually share nearly equally the traits of the parent species.
The Age of Fishes.—The age of fishes is seldom measured by a definite period of years. Most of them grow as long as they live, and apparently live until they fall victims to some stronger species. It is reputed that carp and pike have lived for a century, but the evidence needs verification. Some fishes, as the salmon of the Pacific (Oncorhynchus), have a definite period of growth (usually four years) before spawning. After this act all the individuals die so far as known. In Japan and China the Ice-fish (Salanx), a very long, slender, transparent fish allied to the trout, may possibly be annual in habit, all the individuals perhaps dying in the fall to be reproduced from eggs in the spring. But this alleged habit needs verification.
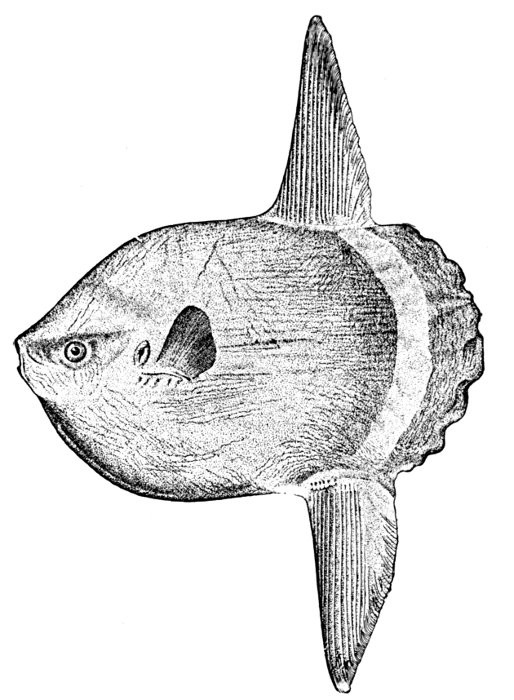
Fig. 111.—Headfish (adult), Mola mola (Linnæus). Virginia.
Tenacity of Life.—Fishes differ greatly in tenacity of life. In general, fishes of the deep seas die at once if brought near the surface. This is due to the reduction of external pressure. The internal pressure forces the stomach out through the mouth and may burst the air-bladder and the large blood-vessels. Marine fishes usually die very soon after being drawn out from the sea.
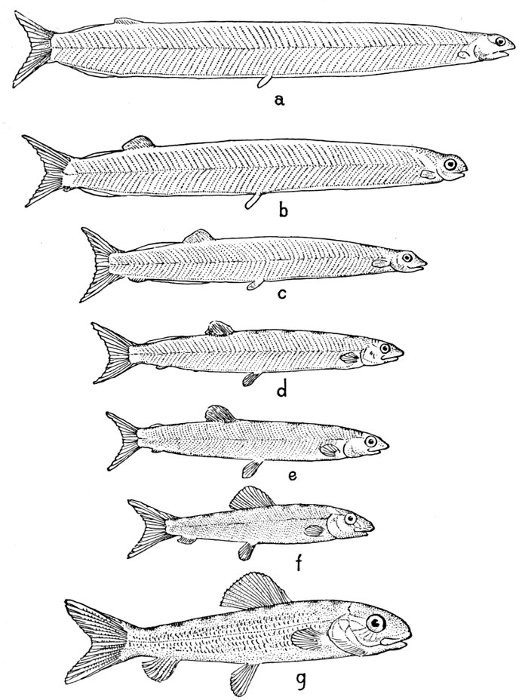
Fig. 112.—Albula vulpes (Linnæus). Transformation of the Ladyfish, from the translucent, loosely compacted larva to the smaller, firm-bodied young. Gulf of California. (After Gilbert.)
Fig. 113.—Development of the Horsehead-fish, Selene vomer (Linnæus). Family Carangidæ. (After Lütken.)
Some fresh-water fishes are very fragile, dying soon in the air, often with injured air-bladder or blood-vessels. They will die even sooner in foul water. Other fishes are extremely tenacious of life. The mud-minnow (Umbra) is sometimes ploughed up in the half-dried mud of Wisconsin prairies. The related Alaskan blackfish (Dallia) has been fed frozen to dogs, escaping alive from their stomachs after being thawed out. Many of the catfishes (Siluridæ) will live after lying half-dried in the dust for hours. The Dipnoan, Lepidosiren, lives in a ball of half-dried mud during the arid season, and certain fishes, mostly Asiatic, belonging to the group Labyrinthici, with accessory breathing organ can long maintain themselves out of water. Among these is the China-fish (Ophiocephalus), often kept alive in the Chinese settlements in California and Hawaii. Some fishes can readily endure prolonged hunger, while others succumb as readily as a bird or a mammal.

Fig. 114.—Ice-fish, Salanx hyalocranius Abbott. Family Salangidæ. Tientsin, China.
Fig. 115.—Alaska Blackfish, Dallia pectoralis (Bean). St. Michaels, Alaska.
The Effects of Temperature on Fish.—The limits of distribution of many fishes are marked by changes in temperature. Few marine fishes can endure any sudden or great change in this regard, although fresh-water fishes adapt themselves to the seasons. I have seen the cutlass-fish (Trichiurus) benumbed with cold off the coast of Florida while the temperature was still above the frost-line. Those fishes which are tenacious of life and little sensitive to changes in climate and food are most successfully acclimatized or domesticated. The Chinese carp (Cyprinus carpio) and the Japanese goldfish (Carassius auratus) have been naturalized in almost all temperate and tropical river basins. Within the limits of clear, cold waters most of the salmon and trout are readily transplanted. But some similar fishes (as the grayling) are very sensitive to the least change in conditions. Most of the catfish (Siluridæ) will thrive in almost any fresh waters except those which are very cold.

Fig. 116.—Snake-headed China-fish, Ophiocephalus barca. India. (After Day.)
Transportation of Fishes.—The eggs of species of salmon, placed in ice to retard their development, have been successfully transplanted to great distances. The quinnat-salmon has been thus transferred from California to Australia. It has been found possible to stock rivers and lakes with desirable species, or to restock those in which the fish-supply has been partly destroyed, through the means of artificially impregnated eggs.
The method still followed is said to be the discovery of J. L. Jacobi of Westphalia (about 1760). This process permits the saving of nearly all the eggs produced by the individuals taken. In a condition of nature very many of these eggs would be left unfertilized, or be destroyed by other animals. Fishes are readily kept in captivity in properly constructed aquaria. Unless injured in capture or transportation, there are few species outside the deep seas which cannot adapt themselves to life in a well-constructed aquarium.
Reproduction of Lost Parts.—Fishes have little power to reproduce lost parts. Only the tips of fleshy structures are thus restored after injury. Sometimes a fish in which the tail has been bitten off will survive the injury. The wound will heal, leaving the animal with a truncate body, fin-rays sometimes arising from the scars.

Fig. 117.—Monstrous Goldfish (bred in Japan), Carassius auratus (Linnæus). (After Günther.)
Monstrosities among Fishes.—Monstrosities are rare among fishes in a state of nature. Two-headed young are frequently seen at salmon-hatcheries, and other abnormally divided or united young are not infrequent. Among domesticated species monstrosities are not infrequent, and sometimes, as in the goldfish, these have been perpetuated to become distinct breeds or races. Goldfishes with telescopic eyes and fantastic fins, and with the green coloration changed to orange, are reared in Japan, and are often seen in other countries. The carp has also been largely modified, the changes taking place chiefly in the scales. Some are naked (leather-carp), others (mirror-carp) have a few large scales arranged in series.
FOOTNOTES:
[9] This account of the normal development of the Teleost fishes is condensed from Dr. Dean's "Fishes Living and Fossil," in which work the details of growth in the Teleost are contrasted with those of other types of fishes.
[10] This paragraph is condensed from Dean's "Fishes Living and Fossil."
CHAPTER XI
INSTINCTS, HABITS, AND ADAPTATIONS

The Habits of Fishes.—The habits of fishes can hardly be summarized in any simple mode of classification. In the usual course of fish-life the egg is laid in the early spring, in water shallower than that in which the parents spend their lives. In most cases it is hatched as the water grows warmer. The eggs of the members of the salmon and cod families are, however, mostly hatched in cooling waters. The young fish gathers with others of its species in little schools, feeds on smaller fishes of other species or of its own, grows and changes until maturity, deposits its eggs, and the cycle of life begins again, while the old fish ultimately dies or is devoured.
Irritability of Animals.—All animals, of whatever degree of organization, show in life the quality of irritability or response to external stimulus. Contact with external things produces some effect on each of them, and this effect is something more than the mere mechanical effect on the matter of which the animal is composed. In the one-celled animals the functions of response to external stimulus are not localized. They are the property of any part of the protoplasm of the body. In the higher or many-celled animals each of these functions is specialized and localized. A certain set of cells is set apart for each function, and each organ or series of cells is released from all functions save its own.
Nerve-cells and Fibres.—In the development of the individual animal certain cells from the primitive external layer or ectoblast of the embryo are set apart to preside over the relations of the creature to its environment. These cells are highly specialized, and while some of them are highly sensitive, others are adapted for carrying or transmitting the stimuli received by the sensitive cells, and still others have the function of receiving sense-impressions and of translating them into impulses of motion. The nerve-cells are receivers of impressions. These are gathered together in nerve-masses or ganglia, the largest of these being known as the brain, the ganglia in general being known as nerve-centres. The nerves are of two classes. The one class, called sensory nerves, extends from the skin or other organ of sensation to the nerve-centre. The nerves of the other class, motor nerves, carry impulses to motion.
The Brain, or Sensorium.—The brain or other nerve-centre sits in darkness, surrounded by a bony protecting box. To this main nerve-centre, or sensorium, come the nerves from all parts of the body that have sensation, the external skin as well as the special organs of sight, hearing, taste, and smell. With these come nerves bearing sensations of pain, temperature, muscular effort—all kinds of sensation which the brain can receive. These nerves are the sole sources of knowledge to any animal organism. Whatever idea its brain may contain must be built up through these nerve-impressions. The aggregate of these impressions constitute the world as the organism knows it. All sensation is related to action. If an organism is not to act, it cannot feel, and the intensity of its feeling is related to its power to act.
Reflex Action.—These impressions brought to the brain by the sensory nerves represent in some degree the facts in the animal's environment. They teach something as to its food or its safety. The power of locomotion is characteristic of animals. If they move, their actions must depend on the indications carried to the nerve-centre from the outside; if they feed on living organisms, they must seek their food; if, as in many cases, other living organisms prey on them, they must bestir themselves to escape. The impulse of hunger on the one hand and of fear on the other are elemental. The sensorium receives an impression that food exists in a certain direction. At once an impulse to motion is sent out from it to the muscles necessary to move the body in that direction. In the higher animals these movements are more rapid and more exact. This is because organs of sense, muscles, nerve-fibres, and the nerve-cells are all alike highly specialized. In the fish the sensation is slow, the muscular response sluggish, but the method remains the same. This is simple reflex action, an impulse from the environment carried to the brain and then unconsciously reflected back as motion. The impulse of fear is of the same nature. Reflex action is in general unconscious, but with animals, as with man, it shades by degrees into conscious action, and into volition or action "done on purpose."
Instinct.—Different animals show differences in method or degree of response to external influences. Fishes will pursue their prey, flee from a threatening motion, or disgorge sand or gravel swallowed with their food. Such peculiarities of different forms of life constitute the basis of instinct.
Instinct is automatic obedience to the demands of conditions external to the nervous system. As these conditions vary with each kind of animal, so must the demands vary, and from this arises the great variety actually seen in the instincts of different animals. As the demands of life become complex, so do the instincts. The greater the stress of environment, the more perfect the automatism, for impulses to safe action are necessarily adequate to the duty they have to perform. If the instinct were inadequate, the species would have become extinct. The fact that its individuals persist shows that they are provided with the instincts necessary to that end. Instinct differs from other allied forms of response to external condition in being hereditary, continuous from generation to generation. This sufficiently distinguishes it from reason, but the line between instinct and reason and other forms of reflex action cannot be sharply drawn.
It is not necessary to consider here the question of the origin of instincts. Some writers regard them as "inherited habits," while others, with apparent justice, doubt if mere habits or voluntary actions repeated till they become a "second nature" ever leave a trace upon heredity. Such investigators regard instinct as the natural survival of those methods of automatic response which were most useful to the life of the animal, the individual having less effective methods of reflex action perishing, leaving no posterity.
Classification of Instincts.—The instincts of fishes may be roughly classified as to their relation to the individual into egoistic and altruistic instincts.
Egoistic instincts are those which concern chiefly the individual animal itself. To this class belong the instincts of feeding, those of self-defense and of strife, the instincts of play, the climatic instincts, and environmental instincts, those which direct the animal's mode of life.
Altruistic instincts are those which relate to parenthood and those which are concerned with the mass of individuals of the same species. The latter may be called the social instincts. In the former class, the instincts of parenthood, may be included the instinct of courtship, reproduction, home-making, nest-building, and care for the young. Most of these are feebly developed among fishes.
The instincts of feeding are primitively simple, growing complex through complex conditions. The fish seizes its prey by direct motion, but the conditions of life modify this simple action to a very great degree.
The instinct of self-defense is even more varied in its manifestations. It may show itself either in the impulse to make war on an intruder or in the desire to flee from its enemies. Among carnivorous forms fierceness of demeanor serves at once in attack and in defense.
Herbivorous fishes, as a rule, make little direct resistance to their enemies, depending rather on swiftness of movement, or in some cases on simple insignificance. To the latter cause the abundance of minnows, anchovies, and other small or feeble fishes may be attributed, for all are the prey of carnivorous fishes, which they far exceed in number.
The instincts of courtship relate chiefly to the male, the female being more or less passive. Among many fishes the male makes himself conspicuous in the breeding season, spreading his fins, intensifying his pigmented colors through muscular tension, all this supposedly to attract the attention of the female. That this purpose is actually accomplished by such display is not, however, easily proved. In the little brooks in spring, male minnows can be found with warts on the nose or head, with crimson pigment on the fins, or blue pigment on the back, or jet-black pigment all over the head, or with varied combination of all these. Their instinct is to display all these to the best advantage, even though the conspicuous hues lead to their own destruction.
The movements of many migratory animals are mainly controlled by the impulse to reproduce. Some pelagic fishes, especially flying fishes and fishes allied to the mackerel, swim long distances to a region favorable for a deposition of spawn. Some species are known only in the waters they make their breeding homes, the individuals being scattered through the wide seas at other times. Many fresh-water fishes, as trout, suckers, etc., forsake the large streams in the spring, ascending the small brooks where they can rear their young in greater safety. Still others, known as anadromous fishes, feed and mature in the sea, but ascend the rivers as the impulse of reproduction grows strong. An account of these is given in a subsequent paragraph.
Fig. 118.—Jaws of Nemichthys avocetta. Jordan and Gilbert.
Variability of Instincts.—When we study instincts of animals with care and in detail, we find that their regularity is much less than has been supposed. There is as much variation in regard to instinct among individuals as there is with regard to other characters of the species. Some power of choice is found in almost every operation of instinct. Even the most machine-like instinct shows some degree of adaptability to new conditions. On the other hand, in no animal does reason show entire freedom from automatism or reflex action. "The fundamental identity of instinct with intelligence," says Dr. Charles O. Whitman, "is shown in their dependence upon the same structural mechanism (the brain and nerves) and in their responsive adaptability."
Adaptation to Environment.—In general food-securing structures are connected with the mouth, or, as in the anglers, are hung as lures above it; spines of offense and defense, electric organs, poison-glands, and the like are used in self-protection; the bright nuptial colors and adornments of the breeding season are doubtfully classed as useful in rivalry; the egg-sacs, nests, and other structures or habits may serve to defend the young, while skinny flaps, sand or weed-like markings, and many other features of mimicry serve as concessions to the environment.
Each kind of fishes has its own ways of life, fitted to the conditions of environment. Some species lie on the bottom, flat, as a flounder, or prone on their lower fins, as a darter or a stone-roller. Some swim freely in the depths, others at the surface of the depths. Some leap out of the water from time to time, as the mullet (Mugil) or the tarpon (Tarpon atlanticus).

Fig. 119.—Catalina Flying Fish, Cypsilurus californicus (Cooper). Santa Barbara.
Flight of Fishes.—Some fishes called the flying-fishes sail through the air with a grasshopper-like motion that closely imitates true flight. The long pectoral fins, wing-like in form, cannot, however, be flapped by the fish, the muscles serving only to expand or fold them. These fishes live in the open sea or open channel, swimming in large schools. The small species fly for a few feet only, the large ones for more than an eighth of a mile. These may rise five to twenty feet above the water.
The flight of one of the largest flying fishes (Cypsilurus californicus) has been carefully studied by Dr. Charles H. Gilbert and the writer. The movements of the fish in the water are extremely rapid. The sole motive power is the action under the water of the strong tail. No force can be acquired while the fish is in the air. On rising from the water the movements of the tail are continued until the whole body is out of the water. When the tail is in motion the pectorals seem in a state of rapid vibration. This is not produced by muscular action on the fins themselves. It is the body of the fish which vibrates, the pectorals projecting farthest having the greatest amplitude of movement. While the tail is in the water the ventral fins are folded. When the action of the tail ceases the pectorals and ventrals are spread out wide and held at rest. They are not used as true wings, but are held out firmly, acting as parachutes, enabling the body to skim through the air. When the fish begins to fall the tail touches the water. As soon as it is in the water it begins its motion, and the body with the pectorals again begins to vibrate. The fish may, by skimming the water, regain motion once or twice, but it finally falls into the water with a splash. While in the air it suggests a large dragon-fly. The motion is very swift, at first in a straight line, but is later deflected in a curve, the direction bearing little or no relation to that of the wind. When a vessel passes through a school of these fishes, they spring up before it, moving in all directions, as grasshoppers in a meadow.
Fig. 120.—Sand-darter, Ammocrypta clara (Jordan and Meek). Des Moines River.
Quiescent Fishes.—Some fishes, as the lancelet, lie buried in the sand all their lives. Others, as the sand-darter (Ammocrypta pellucida) and the hinalea (Julis gaimard), bury themselves in the sand at intervals or to escape from their enemies. Some live in the cavities of tunicates or sponges or holothurians or corals or oysters, often passing their whole lives inside the cavity of one animal. Many others hide themselves in the interstices of kelp or seaweeds. Some eels coil themselves in the crevices of rocks or coral masses, striking at their prey like snakes. Some sea-horses cling by their tails to gulfweed or sea-wrack. Many little fishes (Gobiomorus, Carangus, Psenes) cluster under the stinging tentacles of the Portuguese man-of-war or under ordinary jellyfishes. In the tide-pools, whether rock, coral, or mud, in all regions multitudes of little fishes abound. As these localities are neglected by most collectors, they have proved of late years a most prolific source of new species. The tide-pools of Cuba, Key West, Cape Flattery, Sitka, Unalaska, Monterey, San Diego, Mazatlan, Hilo, Kailua and Waiahæ in Hawaii, Apia and Pago-Pago in Samoa, the present writer has found peculiarly rich in rock-loving forms. Even richer are the pools of the promontories of Japan, Hakodate Head, Misaki, Awa, Izu, Waka, and Kagoshima, where a whole new fish fauna unknown to collectors in markets and sandy bays has been brought to light. Some of these rockfishes are left buried in the rock weeds as the tide flows, lying quietly until it returns. Others cling to the rocks by ventral suckers, while still others depend for their safety on their powers of leaping or on their quickness of their movements in the water. Those of the latter class are often brilliantly colored, but the others mimic closely the algæ or the rocks. Some fishes live in the sea only, some prefer brackish-water. Some are found only in the rivers, and a few pass more or less indiscriminately from one kind of water to another.
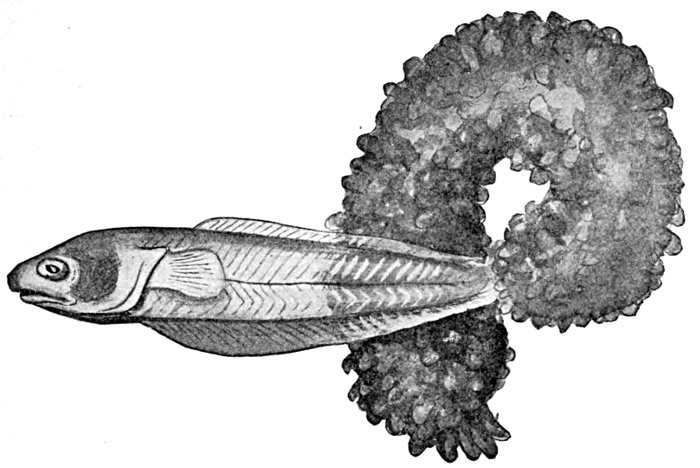
Fig. 121.—Pearl-fish, Fierasfer acus (Linnæus), issuing from a Holothurian. Coast of Italy. (After Emery.)
Fig. 122.—Portuguese Man-of-war Fish, Gobiomorus gronovii. Family Stromateidæ.
Migratory Fishes.—The movements of migratory fishes are mainly controlled by the impulse of reproduction. Some pelagic fishes, especially those of the mackerel and flying-fish families, swim long distances to a region favorable for the deposition of spawn. Others pursue for equal distances the schools of menhaden or other fishes which serve as their prey. Some species are known mainly in the waters they make their breeding homes, as in Cuba, Southern California, Hawaii, or Japan, the individuals being scattered at other times through the wide seas.
Anadromous Fishes.—Many fresh-water fishes, as trout and suckers, forsake the large streams in the spring, ascending the small brooks where their young can be reared in greater safety. Still others, known as anadromous fishes, feed and mature in the sea, but ascend the rivers as the impulse of reproduction grows strong. Among such fishes are the salmon, shad, alewife, sturgeon, and striped bass in American waters. The most remarkable case of the anadromous instinct is found in the king salmon or quinnat (Oncorhynchus tschawytscha) of the Pacific Coast. This great fish spawns in November, at the age of four years and an average weight of twenty-two pounds. In the Columbia River it begins running with the spring freshets in March and April. It spends the whole summer, without feeding, in the ascent of the river. By autumn the individuals have reached the mountain streams of Idaho, greatly changed in appearance, discolored, worn, and distorted. The male is humpbacked, with sunken scales, and greatly enlarged, hooked, bent, or twisted jaws, with enlarged dog-like teeth. On reaching the spawning beds, which may be a thousand miles from the sea in the Columbia, over two thousand in the Yukon, the female deposits her eggs in the gravel of some shallow brook. The male covers them and scrapes the gravel over them. The female salmon does as much as the male in covering the eggs. Then both male and female drift tail foremost helplessly down the stream; none, so far as certainly known, ever survive the reproductive act. The same habits are found in the five other species of salmon in the Pacific, but in most cases the individuals do not start so early nor run so far. The blue-back salmon or redfish, however, does not fall far short in these regards. The salmon of the Atlantic has a similar habit, but the distance traveled is everywhere much less, and most of the hook-jawed males drop down to the sea and survive to repeat the acts of reproduction.
[10] This paragraph is condensed from Dean's "Fishes Living and Fossil."
Development of the Bony Fishes.[9]
The Larval Development of Fishes.[10]



















