

автордың кітабын онлайн тегін оқу Психофизиология. Психологическая физиология с основами физиологической психологии
Е. И. Николаева
Психофизиология. Психологическая физиология с основами физиологической психологии. Учебник
Рекомендовано Министерством образования Российской Федерации в качестве учебника для студентов высших учебных заведений, обучающихся по психологическим и биологическим специальностям
Рецензент доктор медицинских наук Л. И. Афтанас Научный редактор доктор биологических наук В. П. Леутин
© Николаева Е. И., 2008
© ООО «ПЕР СЭ», 2008
© «Логос», 2008
Предисловие
Книга, которая находится у вас в руках, выходит за рамки традиционного учебника по психофизиологии. Ее название свидетельствует о том, что авторский взгляд на соотношение психических и физиологических процессов представляет собой тесное переплетение системного подхода, глубоко проработанного отечественными психофизиологами, с концепциями, предлагаемыми в рамках близкого психофизиологии направления – физиологической психологии. Именно поэтому в учебнике представлены темы, ранее не встречавшиеся в подобного типа изданиях, например психофизиология пола, паранатальная психофизиология, психофизиология аддиктивного поведения и психофизиология старения. Тем не менее введение этих разделов в учебник в настоящее время кажется весьма обоснованным, поскольку современный исследователь изучает не просто психические и физиологические процессы усредненного испытуемого, но рассматривает все эти процессы у мужчины или женщины, находящихся на определенном этапе своего индивидуального развития в соответствующем возрастном периоде.
Другой особенностью учебника является введение достаточного количества спорных теоретических представлений, предложение студенту разных точек зрения на один и тот же вопрос, часто без предоставления окончательного единственно правильного ответа. Это встреча не с устоявшимися окончательными истинами, но с живой психофизиологической наукой, и она, встреча, кажется мне знаковой для нашего времени, поскольку отражает тенденции современного общества стремиться к рассмотрению явлений во всей их многогранности.
И, наконец, бесспорным преимуществом учебника, которое заметно любому, взявшему книгу в руки, является обилие иллюстраций и легкий авторский стиль изложения. Большая часть наших весьма глубоких по содержанию учебников, обращенных к студенту, удручающе бедна наглядным материалом, что крайне затрудняет процесс усвоения, и слишком пересыщена терминологией.
Надеюсь, что всякий, кто откроет первую страницу учебника, не сможет не дочитать его до конца, как это случилось со мной.
Доктор медицинских наук заместитель директора по науке Государственного научно-исследовательского института физиологии СО РАМН

Введение
Психофизиология (психологическая физиология) – наука о физиологических основах психической деятельности человека. Она изучает поведение и внутренний мир индивидуума через призму физиологических изменений.
Практическую психофизиологию каждый человек осваивает с детства, обучаясь сопоставлять собственные переживания и ощущения с теми физиологическими реакциями, которыми они сопровождаются. Точно так же мы пытаемся догадаться о движущих силах поступков других людей, наблюдая за изменением их физиологических реакций. Бытует мнение, что такие реакции в большей мере отражают состояние человека, чем его слова. Разговорный язык фиксирует многочисленные связи между эмоциями и поведением в таких выражениях, как «ноги стали ватными», «это его подкосило», «сердце в пятки ушло» и др.
В «Сказке о мертвой царевне и семи богатырях» А. С. Пушкина, созданной на основе народных сказаний, в большом количестве сохранены подобные ассоциации. Так, встреча царицы с царем передается следующим образом: «Восхищенья не снесла и к обедне умерла».
Исторические факты свидетельствуют о многочисленных попытках анализа психологического состояния человека по его физиологическим реакциям, что и составляло багаж практических знаний по психофизиологии во всех уголках мира. Например, Александр Македонский отбирал в свое войско солдат, резко поднося к лицу новобранца зажженный факел. Если лицо вспыхивало румянцем из-за покраснения кожи, претендент становился солдатом, если же лицо бледнело, то у него не было шансов стать воином.
Сейчас мы объясняем эти реакции дифференцированной активацией при стрессе двух отделов вегетативной нервной системы. Активация симпатической нервной системы, мобилизующей организм на борьбу, связана с приливом крови к мышцам, что фиксируется в покраснении лица. Возбуждение парасимпатической нервной системы, поведенчески реализующееся через замирание или расслабление, сопровождается приливом крови к внутренним органам, что проявляется в побледнении лица. Александр Македонский нуждался в победителях, агрессивно реагирующих в бою, поэтому тест его, физиологический в своей основе, позволял выявлять людей, готовых сражаться до победного конца.
Древние народы использовали знания практической психофизиологии для объективизации решений в трудных ситуациях, хотя многие из них не имели под собой реальной основы. Например, в средневековой Европе считалось, что женщина, весящая менее 49 кг вместе с помелом, непременно должна быть ведьмой (Этинген, 1988). На Руси полагали, что праведный человек, брошенный в воду, утонет, а лживый всплывет. В Китае подозреваемого в преступлении заставляли взять в рот пригоршню риса. Если он мог его выплюнуть, значит был невиновен, поскольку считалось, что у преступника пересыхает во рту и у него не наберется достаточного количества слюны. В действительности же пересыхает во рту не у виновного, а у человека, находящегося в стрессе. Эта вегетативная реакция опосредована адренергическими механизмами, уменьшающими слюноотделение.
Попытку применения психофизиологических знаний в практике Московского уголовного розыска в 30-х годах XX века предпринял А. Р. Лурия. Он использовал ассоциативный тест для выявления преступников среди подозреваемых.
Первые систематические наблюдения о связи изменения частоты пульса с эмоциями провел древнеримский врач Гален. Он описал резкое учащение пульса у женщины в тот момент, когда при ней произносили имя ее возлюбленного (Хэссет, 1981).
В настоящее время психофизиология имеет значительный инструментарий для исследования своего предмета – соотношения психических переживаний и мозговых изменений, их обусловливающих. Накоплено значительное количество данных, позволяющих объяснять и предсказывать те психологические изменения, которые должны сопровождать конкретные физиологические сдвиги. В то же время многое еще остается непознанным.
Распространение психофизиологических знаний за пределы узкого круга специалистов представляется крайне важным: оно позволяет заменить ошибочные представления «народной» психофизиологии на реальные знания, имеющиеся в арсенале науки. Эти знания создают необходимую основу для формирования осознанных представлений о здоровом образе жизни каждого человека.
В названии предлагаемого учебника фигурируют две науки: физиологическая психологияи психологическая физиология. Термин физиологическая психологияввел В. Вундт в конце XIX века. Он использовал его для названия науки, задачей которой явилось изучение психологических возможностей животных в экспериментальных условиях, направленных на изменение состояния их мозга, а также физиологических основ психической деятельности человека. Термин психологическая физиологияпредложил А. Р. Лурия (1978) для выделения системного подхода в исследовании психических процессов, сформировавшегося в Советском Союзе. С точки зрения ученого, физиологическая психология акцентируется на отдельных физиологических процессах, лежащих в основе психических явлений, тогда как центральной задачей психофизиологииявляется анализ целостных форм психической деятельности. Эта наука опиралась на теорию функциональных систем П. К. Анохина (1968). В некоторых источниках психологическую физиологиюсвязывают исключительно с исследованиями психики человека, тогда как физиологическую психологию – с результатами экспериментов на животных. Современные методы, позволяющие неинвазивное (без операционного вмешательства) проведение исследования, постепенно стирают границу в результатах, полученных на человеке и на животных. А потому, рассматривая физиологические процессы, происходящие у человека, мы будем привлекать материалы физиологической психологии, делая выводы более доказательными. Кроме того, современная физиологическая психология так же, как и психологическая физиология, изучает физиологические основы целостного поведения животных и человека.
Появление новых методов исследования стирает рамки не только между психофизиологиейи физиологической психологией. Так, физиология высшей нервной деятельности (ВНД), созданная трудами И. П. Павлова и его учеников, также имеет своей задачей изучение физиологических основ психической деятельности. Термин ВНД введен И. П. Павловым для того, чтобы подчеркнуть отличие собственного подхода в исследовании психики, основанного на эксперименте, от интраспективных психологических методов исследования. Современная психофизиология владеет многочисленными объективными методами получения информации, поэтому нет необходимости выделять физиологию ВДН в отдельную дисциплину.
Другая наука – нейропсихология – сформировалась на стыке нескольких дисциплин (нейрохирургии, психологии, физиологии) и базировалась на анализе последствий локальных поражений головного мозга. Значительный вклад в разработку ее основных положений внес А. Р. Лурия, создавший теорию системной динамической локализации психических процессов. Появление томографии расширяет возможность неинвазивного анализа мозга человека в норме и при патологии, тем самым сближая задачи психофизиологии и нейропсихологии.
Наконец, в настоящее время появилась новая дисциплина – нейронаука, задачей которой является объединение не только данных и подходов перечисленных наук, но и биохимии мозга, нейроиммунологии для создания единой концепции взаимосвязи происходящих в мозге процессов с психической жизнью человека. От успехов этой дисциплины будет зависеть то, насколько долго сохранится отдельная дисциплина психофизиология.
Учебник включает в себя тематически традиционные для такого рода руководств разделы. Отличительной его особенностью является наличие глав, посвященных функциональной асимметрии мозга, паранатальной психофизиологии, психофизиологии пола и аддиктивному поведению. Необходимость введения данного материала обусловлена тем, что исследования в этих направлениях в последнее время достигли больших успехов, но не получили адекватного освещения за пределами узкой аудитории специалистов.
Словарь
Психофизиология —
наука о связи психических переживаний с физиологическими процессами, лежащими в основе этих переживаний, изучает поведение и внутренний мир человека через призму физиологических изменений.
Физиологическая психология —
наука, задачей которой является изучение психологических возможностей животных в экспериментальных условиях, направленных на изменение состояния их мозга. В настоящее время представляет собой науку о физиологических основах психической деятельности человека и животных.
Глава 1
Биологические основы психики
Мозг и психические процессы
Все многообразие взглядов на взаимоотношения между мозгом и психическими процессамиможно свести к трем подходам. Первый предполагает, что психические явления обусловлены исключительно активностью мозга, т. е. за всеми психическими изменениями стоят те или иные процессы, происходящие в мозговой ткани. Предполагается, что не все соотношения этих изменений можно измерить и продемонстрировать в настоящее время, но совершенствование технических возможностей науки приведет в дальнейшем к их обнаружению и пониманию.
Второй подход заключается в том, что только низшие психические процессы связаны с мозгом, тогда как духовная жизнь человека определяется высшим существом (например, Богом). И, наконец, третий подход вытекает из предположения, что мозг является лишь приемником некоторых процессов, которые возникают где-то во вселенной и транслируются через организм человека.
Психофизиология как наука о взаимоотношениях мозгаи психикиполностью опирается на первый подход. Она использует в своем арсенале теоретические позиции, которые построены на фактах, соответствующих требованиям науки в ее современном смысле слова. Это означает, что такие факты могут быть выявлены любым человеком, если он будет для их получения соблюдать соответствующие условия. Эти условия заключаются в том, что ученый работает в рамках определенной парадигмы – принятого на данный момент эталона проведения исследования. Парадигма включает теоретические представления, выведенные на основании предыдущих знаний, методологию, то есть правила проведения исследования, выработанные предыдущими учеными, и инструментарий – оборудование, которое позволяет в той или иной мере ответить на поставленные в исследовании вопросы. Психофизиология, как и любая другая наука, отвечает принципу верифицируемости, то есть любое ее положение должно быть доказано, и принципу фальсифицируемости, то есть любое ее положение может быть опровергнуто в процессе эмпирического исследования. И хотя современная психофизиология способна объяснить лишь те факты, которые связаны с функционированием отдельных нейронов или их цепей, исследователи уверены, что в дальнейшем накопление знаний позволит понять и причину таких явлений, как, скажем, любовь и ненависть (Delgado, 1998).
Проблема соотношения психического и физиологического называется психофизиологической. Она до сих пор не имеет решения. В данном учебнике описаны многочисленные факты взаимного влияния мозга и психики. Тем не менее причинно-следственная связь между ними не поддается описанию на современном уровне состояния науки.
Наиболее простое решение этой проблемы связано со сведением психического к физиологическому, то есть отрицанием специфичности психических процессов. Эту точку зрения, широко распространенную на ранних этапах развития науки, в настоящий момент разделяет узкий круг специалистов.
Другая группа исследователей описывает психические процессы как особые в нервной системе, существенно отличающиеся от остальных, но тем не менее являющиеся нервными.
И, наконец, есть точка зрения, что физиологические и психические процессы имеют различные закономерности, не сводятся друг к другу, хотя психическая активность обусловлена деятельностью мозга. Более подробно психофизиологическая проблема будет рассмотрена в главе 12 «Психофизиология осознанных процессов».
Значительный вклад в обоснование этой точки зрения внес австрийский анатом Франц Иосиф Галль. Он создал науку кефалоскопию, более известную в наше время как френология. Он полагал, что способности людей в той или иной области определяются объемом нервной ткани мозга, которая отвечает за эти способности. Большие скопления нервной ткани, с его точки зрения, деформируют находящуюся над ней черепную кость, вызывая развитие шишек на голове. И хотя сейчас эти представления могут вызывать лишь улыбку, в начале XIX века в магазинах можно было встретить муляжи головы с раскрашенными участками черепа, ответственными за те или иные способности (рис. 1.1).
Выдающийся ученый Чарльз Дарвин чуть было не поплатился своей будущей карьерой из-за пристрастия его современников к легким предсказаниям (по шишкам на голове, линиям на руке и т. д.). Капитан корабля «Бигль» (на котором Дарвин путешествовал, сделал важные открытия, а впоследствии на их основе создал теорию эволюции) увлекался предсказанием способностей людей по рисунку линий на стопе. Стопа ученого явно не внушала ему доверия, и теория эволюции могла бы не возникнуть, если бы капитан, подбирая состав команды для плавания, опирался исключительно на этот метод.
Тем не менее открытия, сделанные Ф. И. Галлем, столь значительны, что он известен в науке как серьезный исследователь. Он обнаружил разницу между строением белого и серого вещества мозга, описал передние и задние корешки спинного мозга и показал, что неразумно приписывать коре мозга функции секреторной железы, как делали до него многие выдающиеся ученые.
Согласно Л. М. Веккеру (2000), существуют основные критические признаки, по которым происходит первичное различение психического акта от физиологического. Они связаны с особенностями отношений между механизмом функционирования органа этого акта и самим актом как результатом этого функционирования. Л. М. Веккер выделяет такие эмпирические признаки, которые отличают психическое от собственно физиологического:
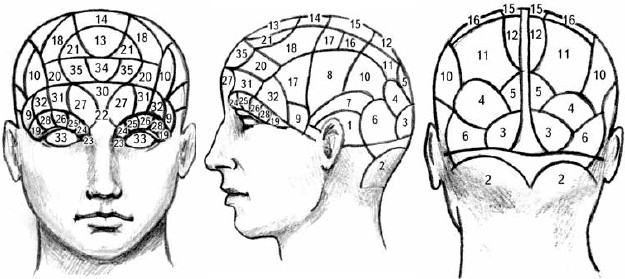
Рис. 1.1.Френологическая карта с обозначением зон мозга, ответственных за различные психические функции.
Эмоции.
I. Склонности.1. Деструктивность. 2. Привлекательность. 3. Чадолюбие. 4. Прилипчивость. 5. Постоянство. 6. Воинственность. 7. Скрытность. 8. Восприимчивость. 9. Конструктивность.
II. Чувства.10. Осторожность. 11. Конформность. 12. Самооценка. 13. Доброжелательность. 14. Благоговение. 15. Непоколебимость. 16. Добросовестность. 17. Надежда. 18. Восхищенность. 19. Идеализирование. 20. Радость. 21. Подражание.
Интеллект
I. Восприятие.22. Индивидуальность. 23. Форма. 24. Размер. 25. Вес и устойчивость. 26. Цвет. 27. Местоположение. 28. Порядок. 29. Счет. 30. Вероятность. 31. Время 32. Гармония. 33. Язык.
II. Рефлексивность.34. Сравнение. 35. Причинность.
1. Предметность. Этот признак состоит в том, что итоговые характеристики любого психического процесса могут быть описаны только в терминах свойств и отношений внешних объектов, физическое существование которых с органом этого психического процесса совершенно не связано и которые составляют его содержание.
Так, восприятие, являющееся функцией органов чувств, нельзя описать иначе, чем в терминах формы, величины, твердости и т. д. воспринимаемого или представляемого объекта. Воспроизведение качеств одного объекта в другом, служащим его моделью, не заключает уникальности психических явлений, поскольку встречается в различных видах и непсихических отображений. Разные предметы – копия и оригинал – могут обладать одной и той же формой, величиной, цветом и т. д. Суть же рассматриваемого исходного критического признака психического процесса заключается в том, что, протекая в своем органе-носителе, этот внутренний процесс в его конечных, результативных параметрах скроен по образцам свойств внешнего объекта.
2. Субъектность. Вторая специфическая особенность психического заключается в том, что в картине психического процесса остаются скрытыми изменения состояния органа-носителя, которые данный процесс реализуют. Итоговые, конечные параметры не могут быть сформулированы на собственно физиологическом языке тех явлений и величин, которые обнаруживаются в мозге. Эта неформулируемость характеристик психических процессов на физиологическом языке внутренних изменений является оборотной стороной их формулируемости лишь на языке свойств и отношений их объекта.
3. Чувственная недоступность. Характеризуется тем, что психические процессы недоступны прямому чувственному наблюдению. Своему носителю-субъекту психический процесс (восприятие или мысль) открывает свойства объекта, оставляя скрытыми изменения в мозге, составляющие механизм этого процесса. Но, с другой стороны, изменения в мозге, открытые с той или иной степенью полноты для стороннего наблюдателя, не раскрывают перед ним характеристик психического процесса другого человека. Человек не воспринимает своих восприятий, но ему непосредственно открывается предметная картина их объектов. Внешнему же наблюдению не открывается ни предметная картина восприятий и мыслей другого человека, ни их собственно психическая «ткань». Непосредственному наблюдению со стороны доступны именно и только процессы в органе, составляющие механизм психического акта.
4. Спонтанная активность. Эта характеристика психического процесса, в отличие от предшествующих, определяет не прямое отношение к объекту или к его непосредственному субстрату – мозгу, а выражение его в поведенческом акте, во внешнем действии, побуждении, которые направляются психическим процессом.
Известно, что вес мозга человека варьирует от 1,5 до 1,8 кг; таким образом, даже в норме разница в весе мозга составляет около 300 г. Еще больший диапазон в весе мозга обнаружен у выдающихся деятелей мировой культуры. Например, вес мозга французского писателя А. Франса составлял около 900 г, тогда как у И. С. Тургенева он был 2 кг 400 г. По некоторым данным, у известного французского естествоиспытателя Л. Пастера было только одно (левое) полушарие мозга, на месте второго имелись лишь зародышевые пузыри (Этинген, 1988). Это свидетельствует о том, что не только вес мозга предопределяет качество психической активности человека.
Среднее количество нейронов– специализированных клеток мозга – составляет 100 млрд, что сопоставимо с числом звезд в системе Млечного Пути. Однако уже подчеркивалось, что сложность строения головного мозга и его функций нельзя объяснить только числом клеток. Все больше подтверждений получает гипотеза, объясняющая взаимосвязь интеллекта не с количеством нейронов, а с числом и особенностью связей между ними. Для понимания этого процесса необходимо ознакомиться со структурой нервной системы и условиями ее работы.
Краткое описание строения нервной системы
В зависимости от поставленных задач научное структурирование нервной системы может происходить различным образом. Каждая из классификаций представляет собой попытку упрощения ее строения для детального изучения. Широко распространена классификация по пространственному признаку, которая позволяет выделять центральную и периферическую нервные системы.
Центральная нервная система (ЦНС)включает структуры, расположенные внутри черепа и позвоночника, – головной и спинной мозг. Все, что находится вне этих костных структур, относится к периферической нервной системе(рис. 1.2).
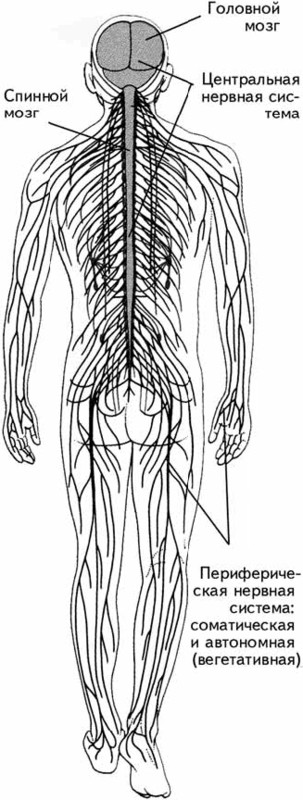
Рис. 1.2.Центральная и периферическая нервные системы.
Головной мозгсостоит из переднего, среднего и заднего (рис. 1.3; табл. 1.1). Передний мозг включает полушария мозга, покрытые корой, миндалину, гиппокамп, базальные ганглии, таламус и гипоталамус. Таламические поля и ядра обеспечивают переключение почти всей сенсорной и моторной информации, входящей и выходящей из переднего мозга. Гипоталамические поля и ядра служат релейными (передаточными) станциями для внутренних регуляторных систем. Средний мозг состоит из крыши среднего мозга, покрышки, четверохолмия, черного вещества. Задний мозг включает варолиев мост, продолговатый мозг, мозжечок. Поля и ядра моста и ствола отвечают за жизненно важную деятельность организма, контролируя дыхание и сердечный ритм. Мозжечок получает и анализирует информацию о положении тела в пространстве.
Таблица 1.1.
Основные разделы головного мозга
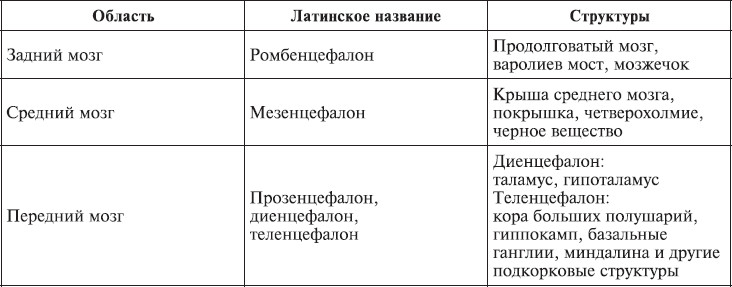
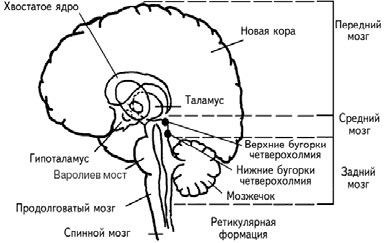
Рис. 1.3.Схематическое расположение подкорковых ядер и структур (Спрингер, Дейч, 1983).
Спинной мозг, который можно рассматривать как продолжение заднего мозга, является центральным коммутатором (переключателем), передающим сообщения из ЦНС на периферию и обратно (рис. 1.4).
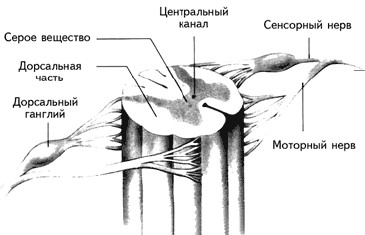
Рис. 1.4.Строение спинного мозга. Дорсальные корешки несут сенсорную информацию в мозг, вентральные корешки – эфферентные команды к мышцам (Kalat, 1992).
Периферическая нервная системасостоит из соматической и вегетативной (автономной). Соматическая нервная системаобеспечивает контроль сокращений поперечно-полосатых мышц, т. е. всей скелетной мускулатуры. Ее нейроны находятся в передних рогах спинного мозга, а их аксоны через передние корешки спинного мозга направляются к скелетным (поперечно-полосатым) мышцам. Там, в области двигательной пластинки мышечного волокна, аксон образует синапс. Соматическая нервная система представлена однонейронным путем (рис. 1.5).
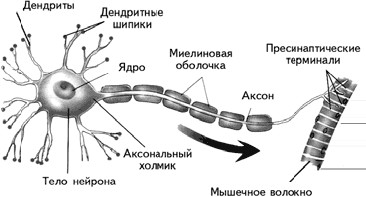
Рис. 1.5.Строение мотонейрона.
Вегетативная нервная системаиннервирует гладкую мускулатуру и управляет деятельностью внутренних органов, поэтому ее еще называют висцеральной. Деление периферической нервной системы на соматическую и вегетативную достаточно условно, поскольку в ЦНС существует значительное перекрытие проекций той и другой, и соматические и вегетативные реакции являются равноправными компонентами любой поведенческой реакции.
Вегетативная нервная система состоит из двух анатомически обособленных систем, являющихся функциональными антагонистами – симпатической и парасимпатической (рис. 1.6). В отличие от соматической нервной системы, имеющей однонейронный путь, пути в вегетативной нервной системе являются двухнейронными. Волокна симпатической нервной системывыходят из грудного и поясничного отделов спинного мозга, где лежит первый симпатический нейрон. Затем они сходятся к симпатическим ганглиям, расположенным вдоль позвоночника, где находится второй симпатический нейрон (рис. 1.6).
Волокна парасимпатической нервной системыначинаются в спинном мозге выше или ниже места выхода симпатических нервов (пара – около, лат.) из черепного и крестцового отдела, а затем сходятся в ганглиях, расположенных не вдоль позвоночного столба, а вблизи от иннервируемого органа (рис. 1.6).
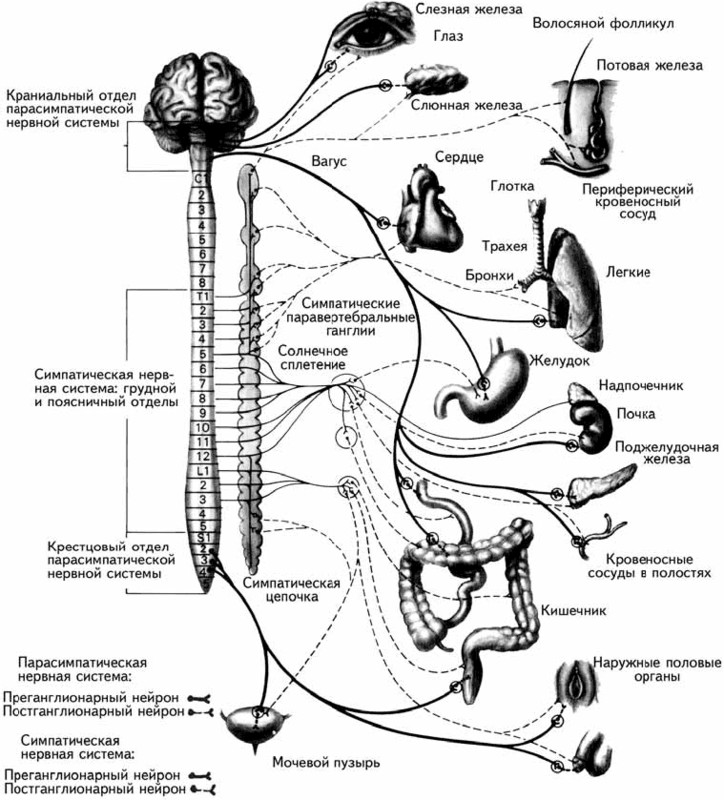
Рис. 1.6.Автономная нервная система и органы, иннервируемые ею (Carlson, 1992).
Особенности расположения ганглиев этих двух систем предполагают различие оказываемого ими эффекта. Действие симпатической нервной системы более диффузно, а парасимпатической – более специфично, поскольку связано только с изменениями в органе, рядом с которым находится ганглий. Эти системы различаются и медиаторами, участвующими в синаптической передаче. Основным медиатором для симпатической нервной системы является адреналин, а для парасимпатической – ацетилхолин.
Клетки мозга
Особенностью живого организма является использование короткоживущих материалов для построения долгоживущих систем. Решение этой на первый взгляд неразрешимой задачи найдено в постоянном обновлении организма. Каждая клетка, каждый орган в нем находятся в состоянии хронического «ремонта», во время которого старые молекулы заменяются новыми. В результате этого структура в целом (например, клетка) живет многие годы, тогда как молекулы вновь и вновь сменяются новыми. Особенно интенсивны эти процессы в мозге, который обновляется на 80 % всего лишь за две недели.
Ведущее значение в деятельности любой клетки принадлежит мембране. В особой мере это относится к нервной системе. В организме человека ее функциями являются интеграцияи коммуникация. Эти функции осуществляются через мембрану: прохождение нервного импульса вдоль аксона, приводящее к возникновению потенциала действия за счет переноса ионов через аксональную мембрану; передача информации от одной клетки к другой представляет собой химические и электрические явления на синаптической мембране; гормональная регуляция связана с восприятием управляющих сигналов – гормонов – через синаптические рецепторы на мембране (Хухо, 1990). Толщина ее составляет в среднем 8 нанометров (нм), что меньше чем 0,00001 мм. В 1934 г. исследователи Дж. Даниэли и Й. Даусон предложили модель, согласно которой клеточная мембрана выглядела как сэндвич (разрезанная булка с маслом внутри). Наружные слои модели составляли белки, а внутри «сэндвича» помещались фосфолипиды (сложные жироподобные молекулы) (Климов, Никульчева, 1995).
Современное представление о структуре мембраны введено С. Зингером и Г. Николсоном, предложившими жидкомозаичную модель мембраны (Singer, Nicolson, 1972). Согласно их гипотезе белки мембраны погружены в гель из двойного слоя фосфолипидов. Эти молекулы имеют два конца, из которых один растворим в воде, а другой нет. Фосфолипиды двух слоев повернуты друг к другу своими нерастворимыми в воде концами (рис. 1.7). Молекулы фосфолипидов, находясь в жидком кристаллическом состоянии, подвижны и могут проникать из слоя в слой (так называемая «флип-флоп диффузия»; flip – щелчок, flop – шлепок, англ.) или меняться местами с соседями (латеральная диффузия). У бактерий одна молекула фосфолипидов завершает полный цикл движения вокруг клетки примерно за 1 сек. – нетрудно представить себе подвижность живой клетки и ее частей (Климов, Никульчева, 1995). Жидкомозаичная модель клеточной мембраны – не более чем модель. Это значит, что она является рабочей гипотезой, в настоящее время наиболее адекватно описывающей мембрану. Любой хорошо запланированный эксперимент может ее изменить или даже отвергнуть (Хухо, 1990). В этом учебнике достаточно часто будет встречаться описание моделей, поэтому следует всегда помнить об относительной надежности этих описаний.
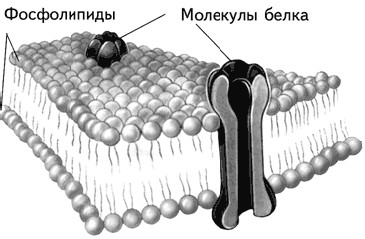
Рис. 1.7.Строение мембраны нейрона (Kalat, 1992).
В каждой клетке тела, кроме клеток крови и генеративных клеток, набор генов одинаков. Однако все разнообразие функционирования клеток определяется набором экспрессирующихся генов (находящихся в активном состоянии, при котором синтезируется продукт, кодируемый данным геном). В каждой клетке эти работающие гены составляют лишь небольшую часть генома. В мозге избирательная генная экспрессия обнаружена в амакриновых клетках в сетчатке, клетках Пуркинье в мозжечке, мотонейронах в спинном мозге.
В процессах метаболизма универсальным источником питания мозговой ткани служит глюкоза. Независимо от того, поступила ли она из кишечника или образовалась в печени, глюкоза с током крови попадает во все ткани организма и используется ими для формирования богатых энергией связей, а также как первичный предшественник углеводов. В клетку глюкозу переносит белок, погруженный в клеточную мембрану. Пять форм такого белка уже достаточно хорошо изучены.
Нейроны
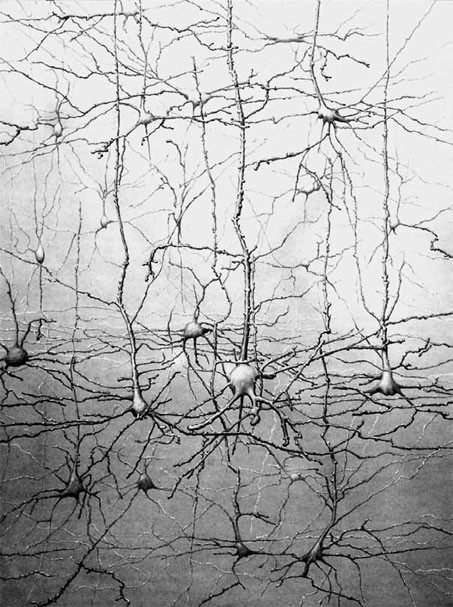
Известны два типа клеток мозга: нейроны и глия. Клеточная теория мозга была сформулирована в 1891 году. Она сменила ретикулярную теорию, согласно которой нервная система представлялась синцитием – гигантской плазматической сетью, не разделенной на отдельные ячейки. Сантьяго Рамон-и-Кахаль, экспериментально подтвердивший клеточную структуру мозга, назвал нейроны «загадочными бабочками души, чьих крыл биение в один прекрасный день – как знать? – прольет свет на тайны психической жизни» (Фишбах, 1992) (рис. 1.8). Человек рождается с окончательным количеством нейронов, не способных к дальнейшему делению при обычных условиях.
С. Рамон-и-Кахаль изучал нейроны, используя метод их фиксации, предложенный Камилло Гольджи. Утверждают, что итальянский врач К. Гольджи открыл этот метод у себя на кухне при свете свечи. Это был метод фиксации клетки двухромовокислым калием и импрегнации серебром (Шеперд, 1987). До К. Гольджи зафиксировать нейроны смог Зигмунд Фрейд. С 1876 по 1881 годы он работал с Эрнстом Брюкке – директором института физиологии при Венском университете, физиологом школы Германа Л. Ф. Гельмгольца. Фрейд предложил метод фиксации нейронов с помощью хлористого золота. Он оказался более дорогостоящим и поэтому менее привлекательным для исследователей.
Рис. 1.8.Фотография нейрона, выполненная Ленарт Нилсон (Kalat, 1992).
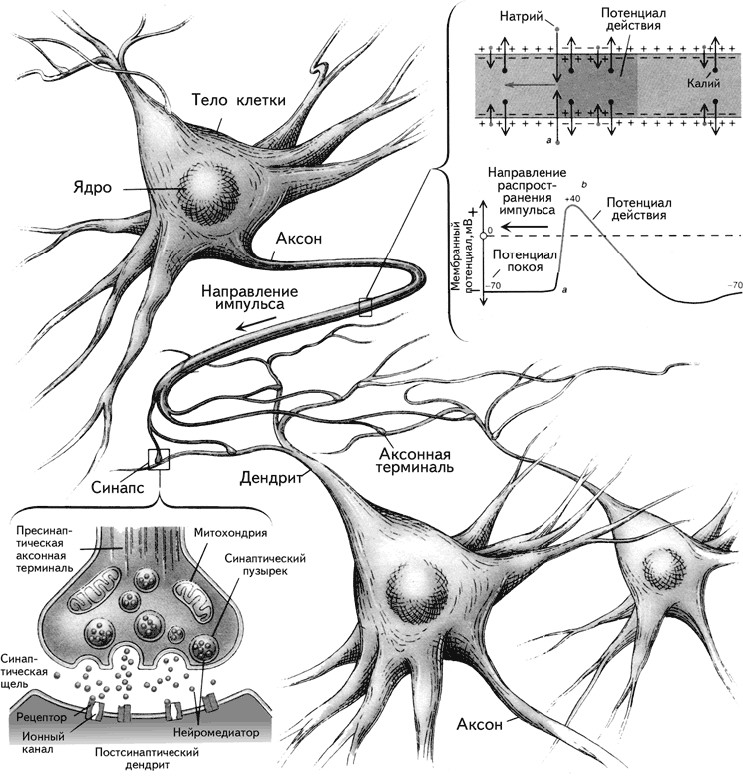
Преимущество метода Гольджи заключалось в том, что серебро, полностью пропитывая нейроны, не проникает в окружающие их глиальные клетки (рис. 1.9). Воспользовавшись этим методом, С. Рамон-и-Кахаль смог увидеть отдельные нейроны и высказал предположение, что мозг образован дискретными единицами. Он впервые описал нейроны как поляризованные клетки, которые с помощью сильно разветвленных многочисленных отростков – дендритов(dendros – дерево, греч.) – получают сигналы и через единственный неразветвленный длинный отросток – аксон (axon – ось, греч.) – посылают информацию другим клеткам (рис. 1.10). Аксон может ветвиться, и его ветви называются коллатералями.В настоящее время доказано, что у нейрона может быть более одного аксона.
Рис. 1.9.Нейроны. Препарат, представленный на рисунке, получен Дж. Роббинс из лаборатории Д. Хьюбела в Медицинской школе Гарвардского университета (Фишбах, 1992)
C. Рамон-и-Кахаль обнаружил фундаментальное различие между клетками с короткими аксонами, взаимодействующими с соседними клетками, и клетками с длинными аксонами, проецирующимися (посылающими сигналы) в другие участки мозга, и продемонстрировал разнообразие нервных клеток (рис. 1.11).
В 1906 г. С. Рамон-и-Кахалю и К. Гольджи была присуждена Нобелевская премия за открытия, сделанные в исследовании структуры мозга. Парадоксально, что С. Рамон-и-Кахаль получил премию за создание клеточной теории мозга, тогда как К. Гольджи, не разделявший эту точку зрения, даже в Нобелевской речи подчеркнул свою уверенность в том, что глия не является клеточной структурой.
Глия
Нейроны составляют лишь 25 % от всех клеток мозга, остальные 75 % клеток относятся к нейроглии(glia – клей, греч.). Это название было дано в 1846 г. Р. Вирховым, полагавшим, что глия – это цементирующая основа для объединения нервных клеток. В среднем глиальные клетки составляют по величине примерно 1/10 размера нейрона. В отличие от нейронов они способны делиться. Именно благодаря им происходит увеличение объема мозга ребенка, составляющего при рождении примерно четверть мозга взрослого. Возникновение опухолей в мозге также связано не с активностью нейронов, а с бесконтрольным делением глиальных клеток.
Глиальные клетки имеют множество функций, но они не передают информацию, как это делают нейроны (рис. 1.12). Мембранный потенциал глиальных клеток выше, чем у нейронов, и определяется разностью концентраций ионов калия во внутри- и внеклеточном пространстве. Это отличает их от нейронов, мембранный потенциал которых формируется как разностью концентраций ионов калия, так и ионов натрия. При возбуждении нейрона из него одновременно выходят ионы К+ и Na+, что ведет к изменению мембранного потенциала расположенных рядом глиальных клеток. Последние частично поглощают ионы калия, функционируя как калиевый буфер, поддерживающий постоянную внеклеточную концентрацию этих ионов. Повышение внеклеточной концентрации калия могло бы снизить порог возбуждения нейрона, что вело бы к его спонтанной активации. Возможно, что именно этот механизм включается при возникновении эпилептических припадков (Хухо, 1990). Выделяют следующие функции глии.
1. Два типа глиальных клеток образуют миелиновую оболочку для аксонов: олигодендроцитыформируют ее в головном и спинном мозге, а Шванновские клетки – в периферической нервной системе. Они обертываются вокруг аксона, изолируя его и ускоряя проведение импульса. Отростки одной глиальной клетки обертываются вокруг разных аксонов, что может способствовать интеграции работы сразу нескольких нейронов.
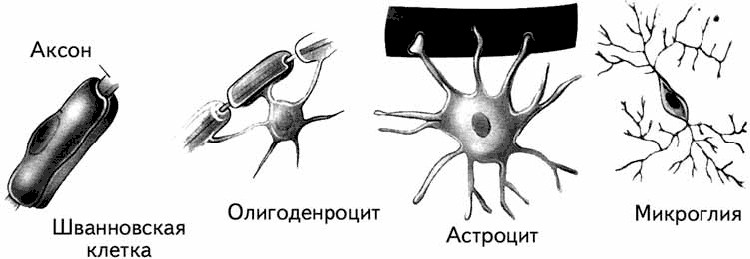
Рис. 1.12.Формы некоторых глиальных клеток.
2. Астроглияи микроглияочищают мозг от погибших нейронов и от ненужного материала, поглощая его.
3. Астроглия также имеет опорную функцию, заполняя промежутки между нейронами и формируя тем самым внутренний скелет.
4. Радиальнаяглия помогает миграции нейронов и направляет аксоны в сторону расположения их мишеней в период эмбрионального развития. Аналогичным образом Шванновские клетки при повреждениях направляют восстанавливающийся аксон к месту иннервации. Они участвуют и в самом востановлении поврежденных нервов. Было показано, что после повреждения аксона Шванновская клетка может заменять утраченное нервное окончание в мышце и даже выделять медиатор (Хухо, 1990). В зрелом мозге радиальная глия перерождается в другие виды глии, осуществляя опорную функцию.
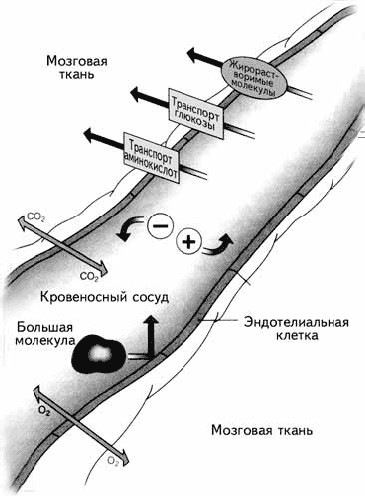
Рис. 1.13.Гематоэнцефалический барьер
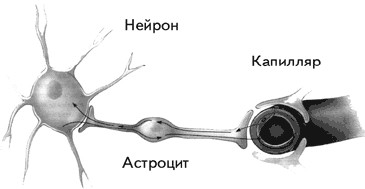
5. Астроглия формирует уникальный защитный слой между нейроном и кровеносным сосудом, так что все вещества из крови могут попасть в нейрон только через глиальную клетку. Этот барьер называется гематоэнцефалическим(haima – кровь, enkephalos – мозг, греч.). Гематоэнцефалический барьер могут преодолевать только маленькие молекулы, например ионы, глюкоза, незаменимые аминокислоты и жирные кислоты (рис. 1.13). Благодаря этому большие молекулы, токсины, вирусы и микробы не могут проникнуть в нейрон, что приводит к значительному повышению толерантности (устойчивости) мозга к вирусным инфекциям.
Существует только одна область мозга, где происходит нарушение гематоэнцефалического барьера, – гипоталамус. В нем находятся клетки, секретирующие либерины и статины, управляющие выделением гормонов из гипофиза. Сосуды непосредственно подходят к секретирующим нейронам, выделяющим свои биологически активные вещества прямо в кровь. Ввиду функциональной необходимости гематоэнцефалический барьер в этом месте нарушается. Гипоталамус можно назвать «ахиллесовой пятой» мозга, поскольку только здесь возможно проникновение инфекций в нервную систему человека.
Наличие гематоэнцефалического барьера при инфекционных поражениях мозговой ткани может препятствовать ее лечению путем введения антибиотиков в кровь. Молекулы лекарства не могут попасть в мозг в нужном количестве и не имеют возможности подойти непосредственно к очагу инфекции. Единственным выходом из этой ситуации остается пункция: лекарство вводится в позвоночный канал, связанный с желудочками мозга, через которые и попадает к очагу инфекции.
Передача информации в ЦНС
Информация в мозге передается по аксонам в виде коротких электрических импульсов, называемых потенциалами действия (ПД). Их амплитуда составляет около 100 мВ, длительность – 1 мс. ПД возникают в результате движения положительно заряженных ионов натрия через клеточную мембрану из внеклеточной жидкости внутрь клетки по специальным натрий-калиевым каналам. Концентрация натрия в межклеточном пространстве в 10 раз больше внутриклеточной.
Существует пассивный и активный транспорт ионов в нейрон. Пассивный (то есть не связанный с расходом энергии) происходит через раздельные Na+ и К+ каналы в мембране аксона. Активный транспорт связан с деятельностью Na+, К+ – насоса, который перекачивает ионы из менее концентрированного раствора в более концентрированный за счет энергии, высвобождаемой при гидролизе АТФ.
В состоянии покоя поддерживается трансмембранная разность потенциалов около 70 мВ (цитоплазма заряжена отрицательно относительно внешней среды). Мембрана практически непроницаема для Na+, тогда как К+ проходит сквозь нее и формирует потенциал покоя. Положительные заряды компенсируются неспособными выйти за пределы клетки анионами. Они и создают суммарный отрицательный заряд. Этот процесс продолжается до тех пор, пока формирующийся внутри клетки отрицательный заряд не будет сдерживать выходящие из нее ионы К+. Устанавливается состояние, при котором число входящих и выходящих ионов К+ уравновешивается.
Несмотря на то, что натрий-калиевый насос выбрасывает ионы натрия из клетки, они очень медленно проникают в клетку. Физическая или химическая стимуляция, деполяризующая мембрану, т. е. снижающая разность потенциалов, увеличивает ее проницаемость для ионов натрия. Поток ионов натрия внутрь клетки еще сильнее деполяризует мембрану (рис. 1.14). Если нейрон возбуждается достаточно интенсивно, то натрий-калиевый насос не успевает предоставить нужное количество натрия для деполяризации, и в этом нейрону помогает глиальная клетка (рис. 1.12).
Синаптическая передача информации
Уже отмечалась важная роль мембраны в передаче информации в мозге. Мембрана представляет собой барьер для прохождения нервного импульса. Именно поэтому связи между нейронами опосредуются химическими передатчиками – нейромедиаторами(mediator – посредник, англ.), выделяющимися из окончаний аксонов в области специализированных межклеточных контактов – синапсов. Синапспредставляет собой мембраны двух соседних нейронов (передающего информацию и получающего ее) и пространство между ними, которое называется синаптической щелью. Синаптическая щель– это пространство шириною около 20 нм между мембранами пресинаптической (мембрана нейрона, находящегося перед синаптической щелью) и постсинаптической (мембрана клетки, находящейся после синаптической щели) клеток (рис. 1.15).
Различают аксо-соматические синапсы, сформированные мембранами аксона и тела (сомы) нейрона, аксо-дендритные, состоящие из мембраны аксона и дендрита, и аксо-аксональные, при которых аксон подходит к другому аксону. Синапс между аксоном и мышечным волокном называется нейромышечной кольцевой пластинкой.
Молекулы медиатора находятся в везикулах– особых пузырьках, расположенных в аксональной терминали (окончании аксона). ПД, достигая аксональной терминали, становится сигналом открытия кальциевых каналов, которые вызывают синхронный эндоцитоз– координированное выделение медиатора из везикул и поступление их в синаптическую щель. Медиатор связывается с рецептором, находящимся на постсинаптической мембране, который инициирует в постсинаптической клетке те или иные изменения в зависимости от вида рецептора. Медиатор, взаимодействуя с рецептором, может способствовать открытию ионных каналов (натрий-калиевых или кальциевых) или через аденилатциклазный механизм активировать внутриклеточного посредника – цАМФ (циклический аденозинмонофосфат) и цГМФ (циклический гуанозинмонофосфат). При открытии натрий-калиевого канала натрий поступает внутрь клетки, что приводит к деполяризации участка мембраны постсинаптического нейрона. Каждый синапс делает лишь незначительный вклад в этот процесс. Однако каждый нейрон непрерывно интегрирует до 1000 синаптических входов, которые суммируются нелинейно (рис. 1.16) и при достижении порогового потенциала вызывают ПД, т. е. распространяющийся вдоль аксона потенциал.
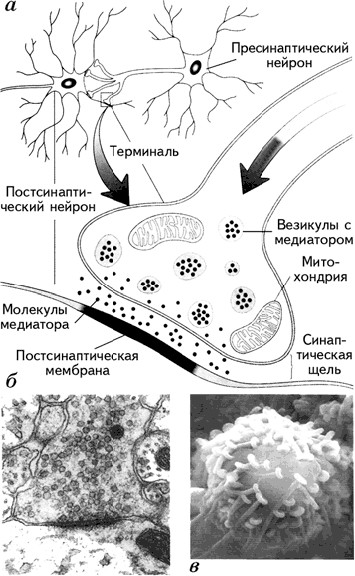
Рис. 1.15.Структура синапса (а); фотография синапса, увеличенного под электронным микроскопом в тысячи раз (выполненная D. D. Kinkel) (б); фотография аксональных терминалей на теле аксона (в) (Kalat, 1992).

Рис. 1.16.Временная и пространственная суммация импульсов (Kalat, 1992).
Синаптическая задержка– время между началом пресинаптической деполяризации и постсинаптической реакцией – составляет 0,5 мс. Всего лишь 1/10 часть этого времени обусловлена диффузией (проникновением) медиатора через синаптическую щель к постсинаптической мембране. Большая же часть времени тратится на открывание Са2+-каналов, через которые кальций проникает в аксонную терминаль и способствует высвобождению медиатора из везикул. Кальций в аксонной терминали имеется в крайне малых количествах. После того как он окажет свое действие, он удаляется, либо связываясь со специальным белком – кальмодулином, либо проникая в эндоплазматический ретикулум. Освобожденный медиатор может действовать как на ауторецепторы (расположенные на пресинаптической мембране), так и на постсинаптические рецепторы.
Согласно принципу английского ученого Г. Дейла, постулирующего метаболическое единство нейрона, во всех окончаниях нейрона выделяется один и тот же медиатор. В настоящее время доказано, что этот принцип касается только пресинаптического единства нейрона. Эффекты, которые вызываются данным медиатором, могут быть различны и зависят от клеток-мишеней (в данном случае постсинаптических нейронов). Знак синаптического действия – повышение постсинаптического потенциала или его падение – определяется не медиатором, а свойствами рецепторов на постсинаптической клетке.
Постсинаптические рецепторы одного пресинаптического нейрона могут фармакологически различаться и контролировать разные ионные каналы. Одна постсинаптическая клетка может иметь более одного типа рецепторов для данного медиатора, и каждый из этих рецепторов контролирует отличный от других механизм ионной проводимости.
Кроме нейронов, суммирующих и передающих информацию к другим клеткам, описаны так называемые пейсмекерные нейроны, способные самостоятельно генерировать электрические импульсы (Alving, 1968). Активность таких нейронов характеризуется синусоидальными колебаниями частотой 0,1–10 Гц и амплитудой 5–10 мВ. Эти нейроны при отсутствии любого внешнего воздействия обеспечивают периодическую генерацию ПД и передачу возбуждения другим нейронам.
Медиаторы
В начале XX века группа английских физиологов, возглавляемая Дж. Лэнгли, показала, что электрическая стимуляция вегетативных нервов вызывает изменения в органах, иннервируемых этими нервами. Оказалось также, что такие изменения можно вызвать инъекцией в организм экстрактов надпочечников. Дж. Лэнгли предположил, что клетки, иннервируемые вегетативными нервами, имеют две рецептивные субстанции – тормозную и возбуждающую.
На основании этих данных Т. Эллиот в 1905 г. выдвинул предположение, что возбуждающие импульсы в вегетативных нервах вызывают выделение адреналина. В 1921 г. австрийский ученый О. Леви обнаружил, что тормозное влияние блуждающего нерва на деятельность сердца опосредуется специфическим веществом, позднее идентифицированным как ацетилхолин. Г. Дейл привел веские аргументы в пользу того, что ацетилхолин является медиатором в вегетативных ганглиях и нервно-мышечных соединениях. Однако доказать наличие синаптической передачи с помощью медиатора, а не электрического потенциала стало возможным только в 50-х годах, когда исследователи начали использовать микроэлектроды и электронный микроскоп.
Все медиаторные соединения – это низкомолекулярные водорастворимые (дипольные) амины или аминокислоты и родственные им вещества. Ацетилхолин и катехоламины синтезируются из циркулирующих в крови предшественников, тогда как аминокислоты и пептиды в конечном счете образуются из глюкозы. Свидетельством консерватизма живой природы является то, что, несмотря на различие циркуляторных систем и метаболических путей, беспозвоночные и позвоночные животные в равной степени используют большинство общих медиаторов (табл. 1.3).
Таблица 1.3.
Характеристика некоторых медиаторов
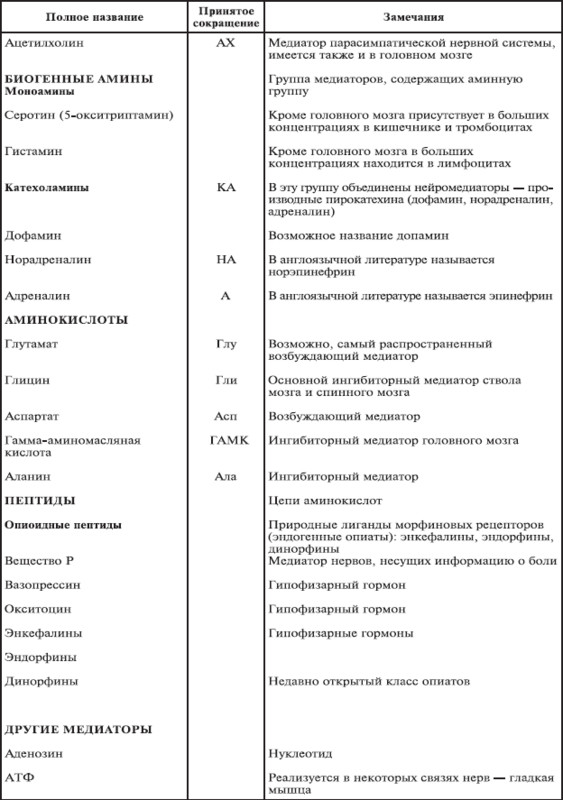
Число пептидов, для которых доказаны медиаторные свойства, постоянно растет. Многие из этих веществ содержат от 2 до 10 аминокислот, что соответствует размеру, с одной стороны, мелких аминокислотных медиаторов, с другой – гормонов. Обилие пептидов создает впечатление неоднородности этой группы веществ. В то же время нарастающая информация о их роли в организме позволяет увидеть универсальные принципы их действия. Предполагается, что нейроэндокринные клетки, секретирующие пептиды, первыми появились в эволюции примитивных нервных систем. По-видимому, нейропептиды, производимые ими, достаточно консервативны, поскольку, как уже упоминалось, одинаковые вещества или близкие последовательности аминокислот обнаружены у филогенетически различных ветвей животных – беспозвоночных и позвоночных. Многие из них найдены не только в мозге, но и, например, в кишечнике. Есть предположение, что все пептид эргические клетки связаны общностью эмбрионального происхождения. Пептиды по сравнению с другими медиаторами оказывают свое действие в чрезвычайно низких концентрациях.
С тех пор как в 1921 г. был идентифицирован первый медиатор, число их в арсенале науки постоянно увеличивается и в настоящее время составляет около 50. Многие биологически активные вещества имеют сходную с ними структуру. Они могут усиливать действие медиаторов (такие вещества называются агонистами) или подавлять их активность (антагонисты). Например, лекарственные препараты, снимающие тревогу (седуксен и др.), усиливают действие тормозного нейромедиатора – гамма-аминомасляной кислоты. Антидепрессанты (например, прозак) являются агонистами серотонина. Кокаин усиливает действие дофамина. Он связывается с белком, удаляющим дофамин из места его активности, тем самым увеличивая время его действия. Никотин активирует рецепторы ацетилхолина. Энкефалины и эндорфины являются природными лигандами морфиновых рецепторов: в норме именно они связываются с рецепторами, с которыми в особых условиях – при употреблении наркотика – взаимодействует морфин.
Важным звеном в нервном пути, который служит мишенью всех наркотиков – амфетамина, никотина, алкоголя и опиатов, – является небольшая часть базальных ганглиев, называемая nucleus accumbens (прилежащее ядро). Антипсихотические препараты (нейролептики) предотвращают связывание дофамина с его рецепторами. Содержащие дофамин нейроны, находящиеся в области вентральной покрышки среднего мозга, посылают свои аксоны в префронтальную кору и базальные ганглии, которые участвуют в двигательном контроле. Это обусловливает побочное действие длительно применяющихся нейролептиков, связанное с развитием дискинезий.
Функции нейрона
В настоящее время можно говорить о наличии трех основных функций нейрона. Наиболее распространенной является суммация возбуждающих и тормозных синаптических потенциалов и передача возбуждения следующему нейрону.
Описаны нейроны (прежде всего нейроны гипоталамуса), обладающие секреторной функцией. Они синтезируют биологически активные вещества – статиныи либерины– и выделяют их в кровеносные сосуды воротной системы гипоталамуса. С током крови эти вещества попадают в переднюю долю гипофиза и способствуют реализации или накоплению его гормонов.
Наконец, существуют нейроны, обладающие спонтанной ауторитмической активностью. Их называют пейсмекерами, или водителями ритма. Эндогенные процессы подобных нейронов приводят к периодическому изменению ионной проницаемости мембраны и генерированию ПД. Взаимодействуя с другими клетками, они синхронизируют активность этих клеток.
Типы нервных волокон
По скорости проведения импульса и строению нервные волокна разделяют на три группы – A, B и C. Волокна типа А делятся на 4 подгруппы: альфа-, бета-, гамма-, дельта-. Альфа-волокна имеют самый большой диаметр (12–22 мк) и обладают наибольшей скоростью проведения возбуждения (70–120 м/сек). Такие волокна проводят информацию к скелетным мышцам и от них в мозг, что позволяет человеку достаточно быстро приспосабливать положение своего тела к ситуации. Остальные нервные волокна имеют меньший диаметр (2–12 мк) и соответственно меньшую скорость проведения импульса. Они несут информацию от сенсорных органов. Гамма-волокна передают возбуждение от моторных нейронов спинного мозга к интрафузальным мышечным волокнам.
Таблица 1.4.
Свойства различных нервных волокон теплокровных (Бабский, 1972).
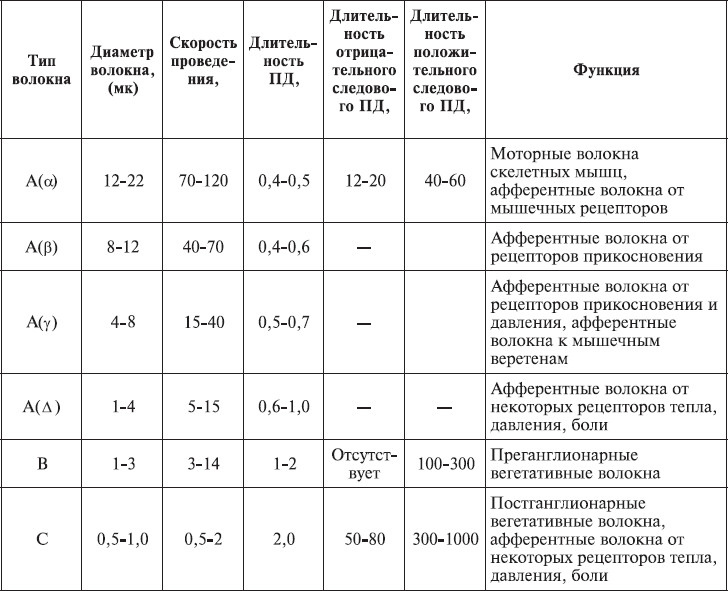
К волокнам типа В относятся миелинизированные волокна, преимущественно преганглионарные, распространенные в автономной нервной системе. Скорость проведения информации по ним составляет 3–14 м/сек.
Волокна типа С – это немиелинизированные волокна, скорость проведения информации в них составляет 2–6 м/сек. Большинство из них являются постганглионарными волокнами автономной нервной системы. Эти волокна проводят также сигналы о боли, тепле, давлении (табл. 1.4) (Бабский, 1972).
Словарь
Вегетативная нервная система—
иннервирует гладкую мускулатуру и управляет деятельностью внутренних органов, поэтому ее также называют висцеральной.
Везикулы—
особые пузырьки, расположенные в аксонной терминали, в которых хранится медиатор.
Гематоэнцефалический барьер—
уникальный защитный слой между нейроном и кровеносным сосудом, образованный глиальными клетками.
Глия—
функционально разнообразные клетки мозга, способные делиться, но не участвующие в процессе передачи информации.
Модуль—
объединение нейронов коры, несущих одну функцию, в виде колонок. Модуль может включать более 100 тыс. клеток, огромное большинство которых образует локальные нейронные сети, выполняющие ту или иную функцию.
Нейрон—
основная функциональная клетка мозга, участвующая в передаче и хранении информации. Это поляризованная клетка, которая с помощью сильно разветвленных многочисленных отростков – дендритов – получает сигналы и через длинный неразветвленный отросток аксон посылает информацию другой клетке.
Периферическая нервная система —
состоит из соматической и вегетативной (автономной).
Соматическая нервная система —
обеспечивает контроль сокращений поперечно-полосатых мышц, т. е. всей скелетной мускулатуры. Ее нейроны находятся в передних рогах спинного мозга, а их аксоны через передние корешки спинного мозга иннервируют скелетные мышцы.
Потенциал действия—
временное изменение мембранного потенциала нейрона под действием приходящих сигналов. Распространяется в обоих направлениях от места возникновения (аксонного холмика).
Синапс—
специализированный межклеточный контакт. Представляет собой мембраны двух соседних нейронов (передающего информацию и получающего ее) и пространство между ними, которое называется синаптической щелью.
Спинной мозг —
можно рассматривать как продолжение заднего мозга; является центральным коммутатором (переключателем), передающим сообщения из ЦНС на периферию и обратно.
Центральная нервная система —
включает структуры, расположенные внутри черепа и позвоночника: головной и спинной мозг. Все, что находится вне этих костных структур, относится к периферической нервной системе.
Контрольные вопросы
1.Что такое психофизиология?
2.В чем единство и различие психофизиологии и физиологической психологии?
3.Структуры центральной нервной системы.
Глава 2
Методы получения психофизиологической информации

Н. Н. Герардов.Плакат театрализованного бала. Музей 10. Художественные собрания СССР: сборник статей. Искусство русского модерна. Сост. А. С. Логинова М.: Советский художник, 1989
Р. У. Эшби (1964) сформулировал положение о том, что сложность объекта исследования предопределяет сложность метода его исследования. Это положение получило название принципа адекватности. Согласно ему, неправомерно изучать сложный объект, каковым является нейрофизиологический механизм психической деятельности, элементарными методами.
Регистрация психофизиологических параметров
По-видимому, самым адекватным прибором для оценки психофизиологических реакций будет другой человек, органы которого в процессе эволюции приспособились для этого. Общаясь, люди ежедневно пользуются этим «прибором», чтобы по поведенческим признакам определить мысли и чувства другого. В то же время каждый человек вносит в свою оценку «шум», зависящий от его воспитания, особенностей развития, наличия или отсутствия болезненных отклонений и так далее. То, что величина такого «шума» у людей различна, мешает использовать этот природный инструмент в качестве экспериментального (рис. 2.1).
Созданные человеком приборы также вносят «шум», но он зависит от конкретных причин, которые можно определить и вычесть из результата, что и делает их более эффективными в исследовании.
Многие физиологические процессы являются по своей сути электрохимическими, поэтому их можно зафиксировать, приложив электроды непосредственно на изучаемый участок тела. Первые доказательства электрической природы физиологических процессов обнаружил Луиджи Гальвани в 1786 г., назвавший эту движущую силу «животным электричеством». Он искренне верил в особые качества этого электричества по сравнению с электричеством, открытым до него физиками (Хэссет, 1981). Дальнейшие исследования не подтвердили его предположения.
Обычно величина регистрируемых физиологических реакций в норме очень невелика, поэтому их усиливают и затем передают на записывающее устройство. Те физиологические процессы, которые имеют отличную от электрической природу, предварительно преобразуют в электрический сигнал, чтобы затем зафиксировать.

Рис. 2.1.Э. Руссо. Ритуалы и мифы нашего времени (обложка к книге E. Imber-Black & J. Roberts, 1992).
Основным записывающим устройством, используемым в психофизиологических исследованиях, является полиграф – прибор, позволяющий одновременно фиксировать изменения электрического потенциала по нескольким каналам (2, 4, 6, 8, 16 и более) (рис. 2.2). Если электроды накладывают на поверхность головы, то получаемая запись называется электроэнцефалограммой (ЭЭГ); если они размещаются на обеих руках, или на руке и ноге, или в области сердца, то запись носит название электрокардиограммы (ЭКГ); в случае, когда они располагаются на тыльной и ладонной поверхности руки, записывается кожно-гальваническая реакция (КГР); при расположении электродов вдоль мышцы можно зафиксировать электрическую активность мышц, т. е. получить электромиограмму (ЭМГ); электроды, расположенные по обе стороны глаз, дают электроокуло грамму (ЭОГ). Более подробно об этих устройствах и методах их применения сказано ниже.
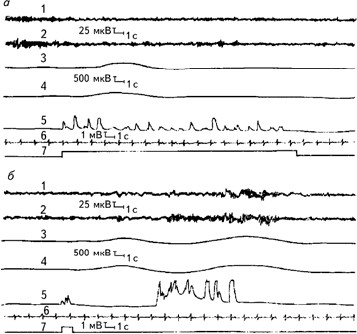
Рис. 2.2.Образец многоканальной записи физиологических функций испытуемого при прослушивании (а) и воспроизведении (б) списка слов. 1, 2 – ЭЭГ, отведения О1 и О2 левое и правое полушария соответственно; 3, 4 – КГР левой и правой руки; 5 – фонограмма (сигналы появляются в моменты предъявления слов магнитофоном или при устных ответах испытуемого); 6 – ЭКГ; 7 – отметка программы опыта (Леутин, Николаева, 1988).
В зависимости от способа наложения электродов и от типа самих электродов при регистрации ЭЭГ, кроме самой ЭЭГ, можно записать также ЭМГ, если человек в процессе обследования будет напрягать лоб или говорить; ЭОГ – если он будет двигать глазами; и даже ЭКГ – если электроды будут, например, сильно прижаты к поверхности кожи и пережмут сосуд, расположенный под ним. Эти ненужные в данный момент для исследователя записи являются помехами и называются артефактами. Электрические сигналы могут генерироваться в некоторых случаях и самим оборудованием, что приводит к появлению наводки. Получение безартефактной записи в психофизиологии сродни искусству и требует многих навыков от исследователя.
Электроэнцефалограмма и методы ее регистрации
В 1929 г. австрийский психиатр Ганс Бергер обнаружил, что с помощью игольчатых платиновых электродов, помещенных на различные точки поверхности головы человека, можно зарегистрировать электрическую активность мозга. Эти записи он и назвал электроэнцефалограммой (ЭЭГ). Хотя ЭЭГ снимается с поверхности головы, Г. Бергер сумел доказать, что часть электрической активности обусловлена деятельностью мозга, а не покрывающих его поверхность тканей (Berger, 1929).
Открытие Г. Бергера было встречено весьма холодно, и этот метод получил признание лишь после того, как Е. Д. Эдриан и Б. Х. Мэттьюз смогли непосредственно продемонстрировать его на заседании Английского физиологического общества в 1935 г. Сам Е. Д. Эдриан был в качестве испытуемого и, закрывая глаза, показал появление альфа-ритма на ЭЭГ (Хэссет, 1981).
Интерпретация электроэнцефалографических данных требует учета целого ряда параметров – формы волн, их амплитуды, расположения электродов, с которых получена запись, и т. д. Наиболее важными из них являются амплитуда волн и частота. Амплитудаизмеряется как расстояние от базовой линии до пика волны. Однако существуют значительные трудности в определении базовой линии, поэтому на практике обычно используют измерение от пика до пика (рис. 2.3). Под частотойпонимается число полных циклов, совершаемых волной за 1 сек. Этот показатель измеряется в герцах (Гц). Величина, обратная частоте, называется периодом волны.Известно, что амплитуда волн ЭЭГ находится в обратно пропорциональной зависимости от их частоты (Cooper e. a., 1980).
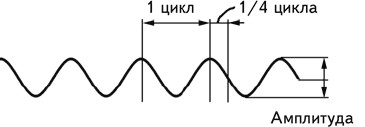
Рис. 2.3.Амплитуда волны и ее цикл.
Ритмичность электроэнцефалографического сигнала позволяет количественно описать записи ЭЭГ. Первым из описанных ритмов электрической активности мозга был альфа-ритм (Berger, 1929). Он представляет собой ритмические колебания электрической активности 8–12 раз в сек (8–12 Гц) с амплитудой около 50 мкВ. Эта активность преобладает у здорового человека в состоянии спокойного бодрствования с закрытыми глазами и наблюдается преимущественно в затылочных областях (рис. 2.4). Альфа-ритм отсутствует у слепорожденных людей (Новикова, 1978). Ритмами той же частоты, но с иной формой волны и в специфических отведениях, являются мю-ритм, регистрируемый в роландовой борозде, каппа-ритм, отмечаемый при наложении электродов в височном отведении (рис. 2.5).
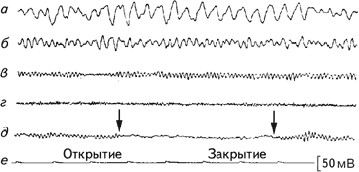
Рис. 2.4.Классификация волн ЭЭГ по частоте. Образцы дельта- (а), тета- (б), альфа- (в), и бета- (г) активности; (д) – блокада альфа-ритма при открытии глаз; (е) – отметка времени (Cooper, 1980).
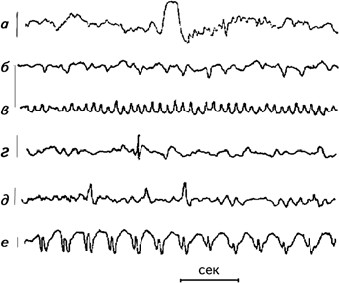
Рис. 2.5.Образцы специфических волн: а– К-комплекс; б– ламбда-волна; в– мю-ритм; г– спайк; д– острая волна; е– комплекс спайк-медленная волна. Слева дается калибровочная линия, соответствующая 100 мВ, справа – расположение электродов на голове. Внизу отметка времени 1 сек (Cooper, 1980).
Г. Бергер описал и другой тип волн – бета-ритм, обнаруживаемый у человека в состоянии активного бодрствования. Это колебания с частотой более 13 Гц и небольшой (около 25 мкВ) амплитудой.
Дальнейшие исследования выявили более медленные колебания, которые были названы тета- и дельта-волнами, а также более быстрые – гамма-волны. Тета-волны имеют частоту от 3,5 до 7,5 Гц и амплитуду от 5 до 100 мкВ, дельта-волны – 1–3,5 Гц и 20–200 мкВ соответственно. Чем медленнее волны, тем больше их амплитуда. Гамма-волны – колебания с частотой более 30 Гц и амплитудой около 2 мкВ (рис. 2.4).
Каждый из ритмов имеет свою преимущественную локализацию: альфа-ритм регистрируется в затылочном и теменном отведениях; тета-волны – в лобных и височных, бета-ритм – в прецентральных и фронтальных, гамма-ритм – в прецентральных, фронтальных, височных, теменных; дельта-ритм не имеет определенной локализации.
При визуальном анализе ЭЭГ обычно определялись два показателя – длительность альфа-ритма и блокада альфа-ритма, которая фиксируется при предъявлении испытуемому того или иного раздражителя (рис. 2.4). Экспериментаторы также пытались обнаружить на ЭЭГ испытуемых особые волны, отличающиеся от фоновой регистрации. Даваемое им название связано либо с их формой, либо с тем местом, где они выявляются. К ним относятся: К-комплекс, ламбда-волны, мю-ритм, спайк, острая волна.
К-комплекс– это сочетание медленной волны с острой волной, вслед за которыми часто идут волны частотой около 14 Гц. К-комплекс возникает во время сна или спонтанно у бодрствующего человека. Максимальная амплитуда отмечается в вертексе и обычно не превышает 200 мВ (Roth e. a., 1956) (рис. 2.5). Ламбда-волны– монофазные положительные острые волны, возникающие в окципитальной области, связанные с движением глаз. Их амплитуда меньше 50 мВ, частота – 12–14 Гц (Green, 1957). Мю-ритм– группа аркообразных или гребневидных волн частотой 7–11 Гц, амплитудой меньше 50 мВ, регистрирующихся в центральных областях головы. Они блокируются двигательной активностью или тактильной стимуляцией (Gastaut e. a., 1952). Спайк– волна, четко отличающаяся от фоновой активности, с выраженным пиком длительностью от 20 до 70 мс. Первичный компонент обычно является негативным (Maulsby, 1971). Острая волна– ясно отличающаяся от фоновой активности волна с подчеркнутым пиком длительностью 70–200 мс. Спайк-медленная волна – последовательность поверхностно негативных медленных волн (частотой 2,5–3,5 Гц), каждая из которых ассоциируется со спайком. Амплитуда этого комплекса может достигать 1000 мВ (Chatrian e. a., 1968) (рис. 2.5).
Визуальный анализ позволяет оценить лишь очевидные изменения биоэлектрической активности мозга (рис. 2.6). В настоящее время разработан метод для регистрации медленных и сверхмедленных потенциалов, имеющих длительность периода от нескольких секунд и более (Илюхина, 1977). Такие потенциалы регистрируются с использованием усилителей постоянного тока (рис. 2.7).
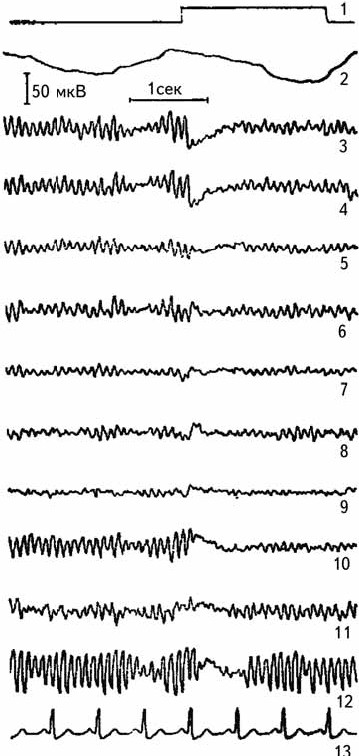
Рис. 2.6.Реакция депрессии альфа-ритма при непрерывном освещении глаза. 1 – отметка раздражителя, 2 – пневмограмма, 3–12 – униполярная регистрация ЭЭГ, 13 – ЭКГ (Лебедева, 1977).
Для более глубокого анализа используются компьютерные методы обработки сигнала, позволяющие извлекать существенно больше информации. Традиционный способ интерпретации электрических сигналов, отраженных в нерегулярных временных рядах, основан на разложении этих сигналов с помощью Фурье-преобразования. При этом в сигнале выделяются частотные компоненты. Затем рассчитывается мощность ЭЭГ в каждой частотной полосе для всех отведений, и с помощью специальных цветовых шкал результаты представляются в виде наглядных картин, где цвет отражает интенсивность ритмов каждого диапазона в различных частях мозга. Этот метод получил название картирования мозга(рис. 2.8). Возможно также использование не цвета, а изолиний при изображении одних и тех же мощностей показателя.
Однако спектральный анализ ЭЭГ в этой парадигме имеет ограниченные возможности в формировании картины электрической активности мозга. В настоящее время появилась возможность применять для анализа ЭЭГ нелинейную теорию систем (Lutzenberger e. a., 1992).
При нелинейном динамическом анализе исследуемые сигналы ЭЭГ рассматриваются как стационарные, то есть предполагается, что значения средних, дисперсии и автоковариационная функция остаются неизменными в течение определенного промежутка времени. Реальные сигналы ЭЭГ сохраняют стационарность в течение длительных интервалов времени с небольшой вероятностью, поэтому эта проблема решается несколькими способами. Прежде всего, для анализа используются относительно короткие временные промежутки (эпохи). Считается, что ошибка в абсолютной оценке параметров за столь короткий срок будет систематической, что позволит корректное сравнение контрольного и экспериментального условий (Pritchard, Duke, 1995). Второй способ решения проблемы заключается в использовании методов, нечувствительных к нестационарности (Skinner, 1994).
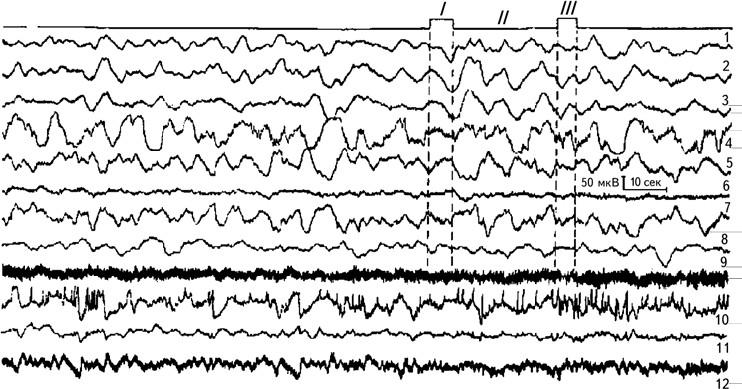
Рис. 2.7.Динамика медленных электрических процессов в подкорковых образованиях головного мозга человека при тесте на оперативную память.
I – предъявление теста; II – этап сохранения; III – воспроизведение теста. 1, 2, 3 – вентролатеральное ядро зрительного бугра; 4 – скорлупа; 5, 6, 7 – бледный шар; 8 – красное ядро; 9 – внутренняя капсула; 10 – передневентральное ядро зрительного бугра; 11 – ретикулярное ядро зрительного бугра; 12 – срединный центр зрительного бугра. Регистрация на энцефалографе с постоянной времени 2,5 сек (Илюхина, 1977).
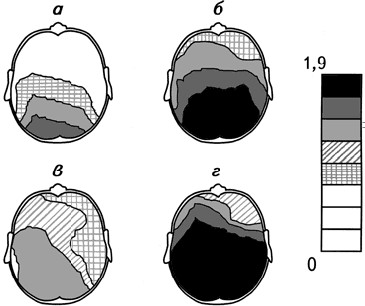
Рис. 2.8.Суммарные топографические карты амплитуды спектра мощности α-ритма ЭЭГ при прослушивании классической (а, б) и рок-музыки (в, г).
а, в – здоровые испытуемые, б, г – больные депрессией. Из каждой карты вычтена соответствующая ей фоновая карта. Справа – шкала амплитуд спектра мощности, мкВ (Михайлова, 1992).
Чтобы отделить сигнала от шума, требуется высокое качество регистрируемого сигнала, либо из кусков качественных записей конструируются наборы суррогатных (искусственных) временных рядов, что позволяет сравнивать результаты нелинейного динамического анализа для оригинальных данных и фильтрованного шума. Несмотря на отсутствие однозначных доказательств действительной хаотичности реального сигнала ЭЭГ, механизм его генерации указывает гипотетически на происхождение от нелинейной динамической системы, управляемой квазипериодическими законами (Афтанас, 2000).
Для характеристики нейрональных динамических систем используются две принципиально различные категории нелинейных динамических мер. К первой категории относится размерностная комплексность(синонимы: корреляционная размерность, фрактальная размерность). Она является показателем общей комплексности (сложности) корковой динамики и отражает ее статические свойства (Pritchard, Duke, 1995). Для изучения динамических характеристик системы, ее изменений во времени, используют энтропию Колмогорова(К2), наибольшую экспоненту Ляпунова(L1) и нелинейное предсказание. Энтропия Колмогорова оценивает степень хаотичности системы, то есть среднюю скорость, с которой информация о системе теряется. Если этот показатель стремится к нулю, то есть нет изменений информации о системе, система становится полностью предсказуемой. Наоборот, для хаотичной системы метрика энтропии имеет определенное конечное положительное значение. И чем больше эта величина, тем более хаотичной является система. Спектр экспонент Ляпунова дает оценку схождения или расхождения близлежащих траекторий в фазовом пространстве, отражая чувствительность системы к исходным значениям. Наличие положительной главной экспоненты является дополнительным свидетельством хаотического поведения изучаемой системы (Афтанас, 2000). Нелинейное предсказание или прогноз характеризует вероятность, с которой может быть предсказана динамика временного ряда. Она основывается на расчете коэффициентов корреляции между наблюдаемым и предсказанными временными рядами. Поэтому она позволяет отслеживать потерю информации во времени.
Впервые оценка фрактальной размерности ЭЭГ произведена в 1985 г. (Babloyantz e.a., 1985). Сейчас получены доказательства относительно более низкой размерности альфа-ритма по сравнению с суммарной ЭЭГ (Pritchard, Duke, 1995).
Динамическая мера фрактальной размерности отражает минимальное количество активированных нейронов и нейрональных цепей, лежащих в основе исследуемого сегмента ЭЭГ. Синхронизация их активности отражает состояние покоя корковых сетей и соответствует высокой степени кооперации между нейронными сетями, что обнаруживается в низкой фрактальной размерности. Десинхронизация, связанная с процессами активной переработки информации в коре головного мозга, характеризует усиление конкурентного взаимодействия между нейронными сетями и сопровождается повышением фрактальной размерности ЭЭГ. Снижение конкуренции между клеточными ансамблями (и, следовательно, снижение уровней фрактальной размерности ЭЭГ) может происходить не только вследствие синхронизации корковых процессов, но и в результате более эффективного использования специализированных для данного типа переработки информации областей коры. В этом случае наблюдается выключение активности нейронных сетей, не вовлеченных в выполнение данного задания, в результате чего более низкие уровни фрактальной размерности ЭЭГ могут отражать интенсификацию процессов переработки информации вовлеченными областями коры.
Другим способом анализа процессов, происходящих в ЦНС, является метод пространственной синхронизации биопотенциалов мозга или когерентности. Этот метод был предложен и разработан М. Н. Ливановым (1972; 1989). Когерентный анализпозволяет определить сходство активности различных областей коры в одном и том же частотном диапазоне и, предположительно, оценить количество корко-корковых соединений и их интенсивность между двумя областями коры (Ливанов, 1989).
Оценка пространственной синхронизации биопотенциалов мозга, основанная на быстром преобразовании Фурье, получила дальнейшее развитие в работах А. М. Иваницкого, предложившего новый метод картирования внутрикорковых взаимодействий. Он основывается на выявлении в ЭЭГ с помощью спектрального анализа пиков активности, совпадающих в разных отведениях по частоте (Иваницкий, 1990, 1996).
В настоящее время разработана новая нелинейная мера – совместная фрактальная размерность, позволяющая количественно оценить степень динамического взаимодействия двух одновременно регистрируемых аттракторов из различных областей коры головного мозга. (Stam e.a., 1996). Каждый из этих аттракторов представляет собой одновременно включенные в процесс самоорганизованные в корковом пространстве нейронные сети, осциллирующие на различных частотах (Stam e.a., 1996). Оценка совместной размерности ЭЭГ не просто отражает фрактальную размерность в области индивидуального отведения, а характеризует степень динамического кооперативного взаимодействия между пространственно разделенными аттракторами. Это принципиально отличает механизмы совместной размерности от процессов, которые отражаются в когерентности, поскольку когерентность оценивает степень взаимодействия дистантно расположенных нейронных сетей, осциллирующих на одной частоте (Ливанов, 1989; Иваницкий, 1996).



















