

автордың кітабын онлайн тегін оқу Эмоциональный мозг
Художник Р. Яцко
Корректор С. Беляева
Павел Симонов
Эмоциональный мозг. — СПб.: Питер, 2021.
ISBN 978-5-4461-3947-7
© ООО Издательство "Питер", 2021
Все права защищены. Никакая часть данной книги не может быть воспроизведена в какой бы то ни было форме без письменного разрешения владельцев авторских прав.
Введение
Эта книга посвящена результатам теоретического и экспериментального исследования эмоций, которое мы вместе с нашими сотрудниками проводили на протяжении последних двух десятилетий.
Интерес к психофизиологии эмоций побудил нас обратиться к анализу творческого наследия К.С. Станиславского. Результатом этого анализа явилась книга «Метод К.С. Станиславского и физиология эмоций», написанная в 1955–1956 гг. и опубликованная издательством АН СССР в 1962 г. Я с благодарностью вспоминаю первого читателя и критика рукописи Леона Абгаровича Орбели. В 1960 г, перейдя в Институт высшей нервной деятельности и нейрофизиологии АН СССР, я получил возможность экспериментальной разработки столь давно увлекавших меня проблем. В тесном научном содружестве с режиссером и театральным педагогом Петром Михайловичем Ершовым мы начали систематическое изучение вегетативных и электрофизиологических сдвигов у актеров в процессе произвольного воспроизведения различных эмоциональных состояний.
И здесь сравнительно скоро мы убедились, что плодотворность такого рода исследований упирается в отсутствие сколько-нибудь разработанной, стройной и обоснованной общей теории эмоций человека и высших млекопитающих животных. Проиллюстрируем наши трудности хотя бы одним примером. Нам не раз приходилось читать о так называемой «эмоциональной памяти». Согласно этим представлениям эмоционально окрашенное событие не только оставляет неизгладимый след в памяти человека, но, став воспоминанием, неизменно вызывает сильнейшую эмоциональную реакцию каждый раз, когда какая-либо ассоциация напомнит о пережитом ранее потрясении. Доверчиво следуя этой аксиоме, мы просили своих исследуемых вспоминать о событиях их жизни, связанных с наиболее сильными эмоциональными переживаниями. Каково же было наше изумление, когда такого рода намеренные воспоминания только в очень ограниченном проценте случаев сопровождались выраженными сдвигами кожных потенциалов, частоты сердцебиений, дыхания, частотно-амплитудных характеристик электроэнцефалограммы. Вместе с тем воспоминания о лицах, встречах, жизненных эпизодах, отнюдь не связанных в анамнезе с какими-либо из ряда вон выходящими переживаниями, подчас вызывали исключительно сильные и стойкие, не поддающиеся угашению при их повторном воспроизведении объективно регистрируемые сдвиги. Более тщательный анализ этой второй категории случаев показал, что эмоциональная окраска воспоминаний зависит не от силы эмоций, пережитых в момент самого события, а от актуальности этих воспоминаний для субъекта в данный момент. Как тут было не вспомнить чеховского Ионыча, который с иронической усмешкой проезжает мимо дома любимой им некогда девушки, мимо балкона, где он провел ночь в состоянии потрясения и восторга. Стало ясно, что дело не в «эмоциональной памяти» и не в эмоциях самих по себе, а в чем-то другом, скрывающемся за фасадом эмоциональных переживаний. Тем более стало ясно, что простое накопление фактов относительно объективных «коррелятов» эмоциональных реакций человека мало что добавит к физиологии эмоций без собственной попытки ответить на вопрос, много раз возникавший в истории науки — на сакраментальный вопрос о том: «что такое эмоция?»
Мы искали ответ на этот вопрос в экспериментах, в литературе, на занятиях семинара молодых сотрудников института, посвященного изучению методологических основ науки о деятельности мозга. Очень важным для нас оказалось общение с физиком Анатолием Никитичем Малышко, который неизменно требовал возможно более точного (пусть рабочего, пусть «для себя»1) определения употребляемых в споре понятий. В феврале 1964 г. появилась «формула эмоций», всесторонней теоретической и экспериментальной разработке которой мы посвятили все последующие годы.
Наш подход к проблеме эмоций целиком принадлежит павловскому направлению в изучении высшей нервной (психической) деятельности мозга. И дело здесь не сводится только к тому, что идея Павлова о формировании и нарушении динамического стереотипа как о критическом звене вовлечения мозгового аппарата эмоций оказалась исходным пунктом «информационной теории эмоций». Может быть, еще важнее общий методологический подход И.П. Павлова к проблеме физиологического и психического при изучении высших форм деятельности мозга.
«Какое было бы основание, — писал Павлов, — как-нибудь различать, отделять друг от друга то, что физиолог называет временной связью, а психолог — ассоциацией? Здесь имеется полное слитие, полное поглощение одного другим, отождествление» [Павлов, 1973, с. 489]. Этот последовательный монизм великого физиолога породил сохраняющийся до сих пор миф об отрицании Павловым психологического аспекта изучения деятельности мозга. Сошлемся на одну из последних статей А.Р. Лурия, где критикуются «многие исследователи в.н. д., стоящие на позиции редукционизма и считающие возможным понимать психические процессы человека как физиологические процессы, построенные по типу условных рефлексов» [Лурия, 1977, с. 68; об этом же — с. 72]. По мнению В.Б. Швыркова, «физиологическая рефлекторная теория давала “чисто физиологическое” объяснение причин и механизмов поведения, в котором отражение мозгом объективной реальности ограничивалось физиологическими процессами, а психика была просто не нужна» [Швырков, 1978, с. 14]. Однако сам Павлов, формулируя свою исследовательскую стратегию, утверждал, что «прежде всего важно понять психологически, а потом уже переводить на физиологический язык» [Павлов, 1954, с. 275]. Если психическое и физиологическое — одно и то же, если «психика не нужна», то откуда же взялись у Павлова эти «прежде» и «потом»? Как можно «переводить на физиологический язык» то, что вообще не существует? В речи на общем собрании XII съезда естествоиспытателей и врачей 28 декабря 1909 г. Иван Петрович Павлов говорил: «Я не отрицаю психологии как познания внутреннего мира человека. Тем менее я склонен отрицать что-нибудь из глубочайших влечений человеческого духа. Здесь и сейчас я только отстаиваю и утверждаю абсолютные, непререкаемые права естественнонаучной мысли всюду и до тех пор, где и покуда она может проявлять свою мощь. А кто знает, где кончается эта возможность?» [Павлов, 1923, с. 88]. Нетрудно видеть, что подлинные взгляды Павлова сильно отличаются от трактовки этих взглядов его комментаторами.
Свой вариант соотношения нейрофизиологического и психического предложил Д.И. Дубровский [1976, 1978], развивая информационный подход к проблеме «сознание и мозг». «Мозговая нейродинамическая система, ответственная за существование образа, — по мнению Дубровского, — есть материальный носитель информации. Связь субъективного образа и соответствующей ему мозговой нейродинамической системы есть связь информации и ее носителя» [Дубровский, 1978, с. 9], причем информация инвариантна по отношению к своему носителю. Два положения, сформулированные Д.И. Дубровским, вызывают протест.
С инвариантностью информации по отношению к носителю трудно согласиться, потому что трудно допустить, чтобы какое-нибудь изменение в содержании психического образа не имело в своей основе соответствующего изменения нейродинамики. Что это за таинственное психическое безразличие к своему нейрофизиологическому «коду»? Современная генетика, откуда заимствованы представления об информации и ее носителе, свидетельствует о том, что любое изменение кода ведет к сильнейшим изменениям содержания наследственной информации при ее «считывании» в процессе онтогенетического развития. Но еще хуже обстоит дело с «мозговым кодом», когда встает вопрос о «реципиенте» этой информации. Д.И. Дубровский не мог уйти от необходимости ответа на подобный вопрос. «Всякое явление сознания носит в принципе отражательный характер и, следовательно, представляет для личности некоторую информацию» [Дубровский, 1978, с. 94. Подчеркнуто нами. — П.С.]. Так вот кто является получателем, читателем информации, закодированной в нейрофизиологических системах, — личность! А что такое «личность» в контексте «информационного подхода» Д.И. Дубровского? Нечто находящееся «над» нейродинамическими системами и закодированной в них информацией? И уж совсем вызывает изумление присущая нам способность «управления некоторым классом мозговых нейродинамических систем своего мозга» [Дубровский, 1978, с. 96]. Кто же это ими управляет? Снова всемогущая «личность»?
Монизм Павлова, ядро его методологии состоит не в том, что он «отрицал» психическое или «сводил» его к нейрофизиологическому. Сила и глубина диалектического мышления Павлова заключается в признании различных аспектов изучения единого по своей объективной природе нейропсихического (высшего нервного) процесса. Психофизиологическая проблема возникла в период, когда деятельность мозга изучали две науки — физиологию и психологию. Создание науки о высшей нервной (психической) деятельности фактом своего рождения ознаменовало первый шаг к диалектическому «снятию» самой проблемы. В истории науки случалось не раз, когда ее развитие вело не к решению вопроса, длительное время волновавшего умы исследователей, но к ликвидации самого вопроса, как неправомерного и бесплодного в свете новых завоеваний человеческого разума.
Наука о высшей нервной деятельности не есть ни физиология, ни психология в традиционном их понимании, ее нельзя однозначно отнести ни к биологическим, ни к социальным наукам, ибо она включает в себя элементы всех этих отраслей знания. Другое дело, что единый процесс отражения объектов и явлений внешнего мира можно рассматривать в различных его аспектах:
• со стороны механизмов этого процесса, то есть как нейрофизиологическое, материальное;
• со стороны его содержания, значения, его отношения к отражаемым объектам внешнего мира и к потребностям субъекта, то есть как психическое, субъективное, идеальное.
Качественная особенность науки о высшей нервной деятельности состоит в том, что она рассматривает оба аспекта отражательной деятельности мозга в их взаимосвязи и взаимообусловленности. В этом, подлинно системном подходе И.П. Павлова к изучению деятельности мозга наиболее полно и ярко воплотились диалектическая сущность и революционный пафос его открытий.
Но мы должны упомянуть еще один аспект, одну ипостась высшей нервной (психической) деятельности мозга, которая явно ускользает от «могучей власти физиологического исследования» и дает основания говорить о суверенности чисто психологического анализа. Суть этого аспекта хорошо сформулировал в одной из своих статей Д.И. Дубровский. «Я не могу исследовать боль как таковую, если не учитываю то фундаментальное обстоятельство, что это явление субъективной реальности другого человека. В противном случае объектом моего исследования будет уже не боль, а нечто иное, скорее всего некоторые объективные изменения в организме человека, сообщившего мне, что он испытывает боль» [Дубровский, 1978, с. 54].
Сделаем этот пример еще более демонстративным с помощью мысленного эксперимента. Представим себе, что на нашей Земле оказался исследователь-инопланетянин, совершенно лишенный чувства боли. В результате своих экспериментов он обнаружил, что усиление разнообразных механических, термических, звуковых и т.п. стимулов в определенный момент вызывает ряд характерных объективно регистрируемых сдвигов в человеческом организме: типичную мимику, речевые реакции (жалобы), повышение кровяного давления и т.д. и т.п. вплоть до изменения активности нервных клеток в определенных отделах головного мозга. Можно ли сказать, что такой высокоинтеллектуальный инопланетянин покинет Землю, поняв, что такое боль как субъективная реальность, как переживание? Разумеется, нельзя. Мы постигаем боль другого исключительно благодаря нашей собственной способности испытывать чувство боли. Никакого иного способа проникнуть во внутренний мир другого человека у нас нет. Та сторона психики, о которой идет речь, лежит за пределами научного познания в общепринятом значении слова «наука».
Другие люди — ничем не заменимое зеркало, благодаря которому субъект не только осознает себя человеком, но и проверяет человечность, всеобщность своего восприятия окружающей действительности. «Лишь отнесясь к человеку Павлу как к себе подобному, человек начинает относиться к самому себе как к человеку. Вместе с тем и Павел как таковой, во всей его павловской телесности, становится для него проявлением рода человек» [К. Маркс и Ф. Энгельс, Соч., 2-е изд., т. 23, с. 62]. Постижение природы человека, а следовательно, и своей собственной природы может носить характер научного познания во всех его разновидностях (философия, психология, антропология, социология, физиология и т.д.). Но отражательная деятельность мозга не исчерпывается знанием, добываемым наукой. Существует и второй путь, значение которого в развитии цивилизации подчас остается как бы в тени триумфальных завоеваний научной мысли. Рядом с «co-знанием» функционирует «со-переживание».
Изучая человеческий мозг, наука имеет дело с коррелятами (речевыми, электрофизиологическими, биохимическими) психических процессов, но для нее остается недоступна их субъективная сторона. Методы науки не в состоянии познакомить нас с переживанием боли, удовольствия, радости, отчаяния и т.п. другого человека. Эту возможность дает только сопереживание, роль которого до сих пор в полной мере не оценена ни теорией, ни практикой воспитания.
Было бы ошибкой полагать, что функции механизма сопереживания ограничиваются одним лишь приобщением нас к внутреннему миру других людей. Представление о сопереживании как о чем-то архаичном, грубом, приблизительном по сравнению с мышлением и имея в виду его изощренную логику, право же несправедливо: вчувствоваться можно не менее глубоко, чем вдуматься. Мир переживаний, сопровождающих процесс общения между людьми, может быть чрезвычайно богат, усложнен и тонок, оставаясь невербализуемым. Достаточно вспомнить об эмоциях, возникающих при восприятии произведений искусства. Хотя одно и то же произведение вызывает у каждого зрителя свой ряд эмоций, в них есть нечто общее, то есть момент сопереживания. В противном случае каждое произведение имело бы очень узкий круг почитателей, а не служило миллионам людей на протяжении веков.
Двойственная природа психики, зависимость процесса отражения от объекта отражения и наделенного потребностями субъекта породили две основные разновидности, две ветви человеческой культуры, взаимно дополняющие друг друга: науку и искусство. Субъективная сторона внутреннего мира личности не является предметом нейрофизиологии, поскольку она не является предметом науки вообще. Отступая от преследующих его по пятам смежных дисциплин — нейрофизиологии, этологии, антропологии, социологии и т.п., психолог в определенный момент оказывается на территории, где он чувствует себя недостигаемым для представителей этих отраслей знания. С облегчением он осматривается вокруг и обнаруживает, что находится на территории... искусства.
Нам было важно остановиться на общеметодологических вопросах для того, чтобы сразу же недвусмысленно определить свой подход к проблеме эмоций. Информационная теория эмоций, которой будет посвящено все последующее изложение, не является ни только «физиологической», ни только «психологической», ни тем более «кибернетической». Она неразрывно связана с павловским системным по своему характеру подходом к изучению высшей нервной (психической) деятельности. Это означает, что теория, если она верна, должна быть в равной мере продуктивна и для анализа явлений, относимых к психологии эмоций, и при изучении мозговых механизмов эмоциональных реакций человека и животных. Ее истинность или ложность должны быть доступны проверке как в психологических, так и в нейрофизиологических экспериментах. Наконец, она должна находить подтверждение в педагогической и клинической практике, в художественных произведениях, посвященных описанию внутреннего мира человека.
В заключение я хочу поблагодарить своих товарищей по работе, чей труд наполнил конкретным фактическим содержанием те или иные разделы экспериментальной разработки интересовавших нас вопросов: И.И. Вайнштейн, М.Н. Валуеву (Русалову), И.Н. Грызлову, А.П. Ершову, А.А. Заничеву, И.С. Иванова, К. Изарда, А.Н. Лукьянова, В.А. Маркевича, А.Я. Мехедову, Н.Г. Михайлову, Р.А. Павлыгину, Д.И. Пайкина, М.Л. Пигареву, Т.Г. Пименову, Л.А. Преображенскую, В.А. Пучкова, К.Ю. Саркисову, О.А. Сидорову, С.Е. Скорикову, С.И. Табачникову, В.Л. Таубкина, В.Д. Труша, М.В. Фролова, С.Н. Чугунову. Многие наши эксперименты были поставлены с участием тех, чьи энтузиазм и время согласно традиции скрываются за инициалами в протоколах наших опытов. Спасибо им всем — актерам высшей квалификации и учащимся театральных учебных заведений, спортсменам-парашютистам и диспетчерам аэропортов, студентам и лаборантам. Их эмоции, записанные перьями регистрирующих приборов остались на страницах наших книг, диссертаций и статей.
С особым волнением и безграничной благодарностью я произношу имена своих учителей — Василия Львовича Симонова и Эзраса Асратовича Асратяна.
Что такое эмоция?
«Путь определений и классификации, который проделывала психология на протяжении нескольких столетий, привел к тому, что психология чувств оказалась самой бесплодной и скучной из всех глав этой науки» [Выготский, 1970, с. 127]. Определения термина «эмоция», которые, казалось бы, должны в наиболее концентрированном виде отражать степень нашего проникновения в природу и внутреннюю структуру определяемого явления, как правило, носят абстрактно-описательный характер или требуют дополнительных разъяснений. Приведем несколько примеров подобных определений для того, чтобы показать причину той неудовлетворенности, которая побудила нас искать собственный ответ на вопрос: «что такое эмоция?».
«Эмоции — одна из важнейших сторон психических процессов, характеризующая переживание человеком действительности. Эмоции представляют интегральное выражение измененного тонуса нервнопсихической деятельности, отражающееся на всех сторонах психики и организма человека» [Лебединский, Мясищев, 1966, с. 222]. Определение эмоции как переживания (а переживания — как эмоции) можно встретить не только у психологов, но и в физиологических работах: «Эмоции — физиологические состояния организма, имеющие ярко выраженную субъективную окраску и охватывающие все виды чувствований и переживаний человека — от глубоко травмирующих страданий до высоких форм радости и социального жизнеощущения» [Анохин, 1964, с. 339]. Правда, физиологический подход к определению эмоций, как правило, хотя и далеко не у всех исследователей, обнаруживает тесную связь эмоций с потребностями организма. «С точки зрения физиологической перед нами стоит задача раскрыть механизм тех конкретных процессов, которые в конечном итоге приводят к возникновению и отрицательного (потребность) и положительного (удовлетворение) эмоционального состояния» [Анохин, 1964, с. 355].
Связь эмоций с потребностями бесспорна, однако считать эмоцию функцией одной лишь потребности вряд ли правомерно. Неудовлетворенная потребность необходима для положительных эмоций не менее, чем для отрицательных. Для того чтобы убедиться в этом, достаточно представить себе состояние человека, который после обильной трапезы получает приглашение снова сесть за стол. Тем не менее гомеостатическое отождествление эмоций с самим фактом возникновения или устранения потребностей (с редукцией драйва) продолжает кочевать по страницам научных работ. «Снижение или устранение мотиваций, — пишет Ю.А. Макаренко [1972, с. 224], — оценивается как нечто приятное, а увеличение мотиваций — как нечто неприятное. Таким образом, мотивации всегда специфичны, почти всегда отрицательны (поскольку отражают наличие потребности) и всегда цикличны».
Наряду с потребностью в определениях эмоций появляется фактор саморегуляции. Согласно Г.X. Шингарову, эмоции есть форма «отражения действительности, сущность (!) которой заключается в саморегуляции функций организма, согласно требованиям условий внешнего мира» [Шингаров, 1970, с. 7]. Но так называемая «саморегуляция функций» — понятие неизмеримо более широкое, чем эмоции. Огромная масса процессов саморегуляции, протекающих в живом организме, не сопровождается какими-либо эмоциональными переживаниями. И Г.X. Шингаров сам делает следующий шаг к растворению эмоций в многообразных проявлениях жизнедеятельности: «...любой психический процесс протекает по принципу: причина — реакция — подкрепление (как «снятие» причины). Какое место занимают эмоции в этой принципиальной схеме деятельности нервной системы? Они существуют на всех этапах этой деятельности» [Шингаров, 1971, с. 137].
Наряду с уже знакомыми нам категориями потребности и регулирования в определениях эмоций появляется фактор отношения, значимости, смысла. «Эмоции — особый класс психических процессов и состояний, связанных с инстинктами, потребностями и мотивами. Эмоции выполняют функцию регулирования активности субъекта путем отражения значимости внешних и внутренних ситуаций для осуществления его жизнедеятельности» [Леонтьев, 1970, с. 553]. С появлением категории значимости (отношение, оценка, смысл, «значимое переживание», «личностный смысл» и другие вариации на ту же тему) определения эмоций становятся удивительно похожи друг на друга независимо от того, кому они принадлежат: философу, психологу, физиологу, фармакологу и т.д. Эмоции есть «деятельность оценивания поступающей в мозг информации о внешнем и внутреннем мире, которую ощущения и восприятия кодируют в форме его субъективных образов» [Додонов, 1978, с. 29]. «Эмоциональные процессы — это специфический класс процессов психической регуляции, приводимых в действие факторами, значимыми для индивида» [Рейковский, 1979, с. 354]. «Эмоция — это такая форма отражательной психической деятельности, где на первый план выступает отношение к окружающей информации, где сигналы информации преобразуются в личностном плане» [Вальдман, 1978, с. 132]. Здесь что ни слово, то вопрос, требующий дополнительных разъяснений: а что такое «отношение», «личностный план» и каким образом сигналы информации «преобразуются в личностном плане»?
«Эмоции — как форма отражения биологического качества раздражителей, его полезности или вредности для организма, входя в функциональную систему поведенческого роста, способны в значительной мере модулировать его направленность и конечный результат» [Вальдман, Звартау, Козловская, 1976, с. 161–162]. Утверждение о том, что полезное для организма оценивается им как «приятное, эмоционально положительное», а вредное — как «неприятное, эмоционально отрицательное», тривиально. Представим на минуту, что сталось бы с эволюцией, если бы живые существа стремились ко всему вредному, разрушительному и избегали все полезное, необходимое для жизни. Сказать, что полезное — приятно, а вредное — неприятно, значит ничего не добавить к самоочевидному положению вещей.
Мы закончим наш лингвистический экскурс определением термина «эмоции» в 3-м (последнем) издании «Большой советской энциклопедии»: «Эмоции... — субъективные реакции человека и животных на воздействие внутренних и внешних раздражителей, проявляющиеся в виде удовольствия или неудовольствия, радости, страха и т.д. Сопровождая практически любые проявления жизнедеятельности организма, эмоции отражают в форме непосредственного переживания значимость (смысл) явлений и ситуаций и служат одним из главных механизмов внутренней регуляции психической деятельности и поведения, направленных на удовлетворение актуальных потребностей (мотивации)» [Леонтьев, Судаков, 1978, с. 169]. Подобное определение можно признать справедливым в самых общих его чертах, однако оно требует уточнений, без которых рискует остаться слишком расплывчатой фразой. Чем определяется значимость (смысл) явлений для субъекта? Почему эта значимость может быть исключительно велика в одном случае или практически отсутствовать в другом? Каким образом значимость и смысл связаны с потребностями и поведением? Ответы на поставленные нами вопросы представляют суть решения проблемы эмоций как специфической формы отражения окружающей действительности.
Впрочем, функция отражения признается за эмоциями далеко не всеми авторами. «При исследовании познавательных процессов, — пишет В.К. Вилюнас в предисловии к русскому переводу книги Я. Рейковского [1979, с. 7], — обычно существует возможность опираться на два ряда явлений: объективный и субъективный, отражаемый и отраженный. По отношению к субъективному отражению первый ряд может служить своего рода образцом, «эталоном» того, что, например, должно или могло бы быть воспринято, запечатлено, заучено, постигнуто мышлением и т.п. ... При исследовании же эмоций такой возможности не существует. Эмоции выполняют функцию не отражения объективных явлений, а выражения субъективных к ним отношений... Поэтому данные о том или ином эмоциональном переживании мы можем сравнивать лишь с данными о других эмоциональных переживаниях у одного и того же человека или у других людей, а не с некоторым объективным «эталоном». Итак, эмоции есть «выражение без отражения», их допустимо сравнивать не с объективными факторами, эти эмоции порождающими и детерминирующими, а лишь с другими эмоциями. Далеко же продвинулась психология чувств за время, прошедшее с тех пор, когда Л.С. Выготский произнес свою грустную сентенцию!
Не найдя ответа на сформулированные выше вопросы, мы обратились к научному наследию И.П. Павлова. В трудах Павлова мы находим указания на два фактора, неразрывно связанные с вовлечением мозговых механизмов эмоций. Во-первых, это присущие организму потребности, влечения, отождествлявшиеся Павловым с врожденными (безусловными) рефлексами. «Кто отделил бы, — писал Павлов, — в безусловных сложнейших рефлексах (инстинктах) физиологическое соматическое от психического, то есть от переживаний могучих эмоций голода, полового влечения, гнева и т.д.?» [Павлов, 1951, с. 335]. Однако Павлов понимал, что бесконечное многообразие мира человеческих эмоций не может быть сведено к набору врожденных (даже «сложнейших», даже жизненно важных) безусловных рефлексов. Более того, именно Павлов открыл тот ключевой механизм, благодаря которому в процесс условнорефлекторной деятельности (поведения) высших животных и человека вовлекается мозговой аппарат, ответственный за формирование и реализацию эмоций. О своем открытии Павлов впервые сообщил 24 августа 1932 г. на X Международном психологическом конгрессе в Копенгагене. Непосредственным поводом для обращения Павлова к проблеме эмоций послужили эксперименты Э.А. Асратяна и ряда других сотрудников павловских лабораторий, посвященные явлению системности (термин Э.А. Асратяна) или динамической стереотипии (термин И.П. Павлова) в работе больших полушарий головного мозга.
На основании этих опытов Павлов пришел к выводу о том, что под влиянием внешнего стереотипа повторяющихся воздействий в коре больших полушарий формируется устойчивая система внутренних нервных процессов, причем «образование, установка динамического стереотипа есть нервный труд чрезвычайно различной напряженности, смотря, конечно, по сложности системы раздражителей, с одной стороны, и по индивидуальности и состоянию животного, с другой» [Павлов, 1973, с. 429]. Не меньший, а иногда еще более напряженный «умственный» (выражение Павлова) труд представляет перестройка сложившегося стереотипа и замена его новым. По мнению Павлова, «описанные физиологические процессы в больших полушариях отвечают тому, что мы субъективно в себе обыкновенно называем чувствами в общей форме положительных и отрицательных чувств и в огромном ряде оттенков и вариаций, благодаря или комбинированию их, или различной напряженности. Здесь чувство трудности и легкости, бодрости и усталости, удовлетворенности и огорчения, радости, торжества и отчаяния и т.д.» [Павлов, 1973, с. 431].
Неделю спустя, 2 сентября 1932 г. Павлов вернулся к этой теме в своем докладе на XIV Международном физиологическом конгрессе в Риме. «Нужно думать, — говорил Павлов с трибуны конгресса, — что нервные процессы полушарий при установке и поддержке динамического стереотипа есть то, что обыкновенно называется чувствами в их двух основных категориях — положительной и отрицательной, и в их огромной градации интенсивностей. Процессы установки стереотипа, довершения установки, поддержки стереотипа и нарушений его и есть субъективно разнообразные положительные и отрицательные чувства, что всегда и было видно в двигательных реакциях животного» [Павлов, 1973, с. 423].
Эту павловскую идею несовпадения (рассогласования — скажем мы сегодня) заготовленного мозгом внутреннего стереотипа с изменившимся внешним мы не раз встретим в той или иной модификации у ряда авторов, обращавшихся к изучению эмоций. По мнению Хоуджа [Hodge, 1935], эмоции возникают в момент, когда высшие мозговые центры не могут обеспечить адекватный ответ на воспринимаемую ситуацию или когда существуют сомнения, колебания относительно возможности успешного ответа. По Хоуджу, сила эмоциональной реакции обратно пропорциональная возможности высших центров мозга адекватно ответить на данную ситуацию. Эмоции представляют неудачу интеграции на уровне церебральной коры.
Близкие теоретические представления были позднее развиты Хеббом [Hebb, 1946] на примере активации врожденного механизма страха. Согласно Хеббу, этот механизм вовлекается в процесс поведения, когда ситуация оказывается «странной» — частично знакомой, частично — нет — и не вполне понятной. Реакция страха основывается скорее на переживании рассогласования, чем на собственно сенсорном восприятии сложившейся обстановки. Результаты систематических экспериментов с разрушением различных структур, относящихся к так называемой лимбической системе, позволили Арнольд [Arnold, 1960] утверждать, что эмоции возникают под влиянием активирующих команд из новой коры, где происходит «сплав ожидания с сенсорным представительством оценки ситуации».
«Биологическая теория эмоций» П.К. Анохина [1964] непосредственно связана с его более общей теорией функциональной системы поведенческого акта. Согласно Анохину нервный аппарат отрицательных и положительных эмоций активируется в тот момент, когда обнаруживается рассогласование или совпадение акцептора действий (афферентной модели ожидаемых результатов) с импульсацией, сигнализирующей о реально достигнутом эффекте. Мы завершим обзор этой линии исследований, органически близких идее И.П. Павлова, сформулированной им в 1932 г., ссылкой на статью К. Прибрама «Новая биология и неврология эмоций. Структурный подход», появившуюся в 1967 г. уже после того, как была опубликована [Симонов, 1964] и доложена на XVIII Международном психологическом конгрессе в Москве «информационная теория эмоций» [Симонов, 1966]. Отметив значение экспериментов Хебба и Линдсли [Lindsley, 1951], показавших, что степень эмоционального напряжения можно количественно определить по его выходу на эффекторные органы, Прибрам заключает: «Изменения вегетативных функций могут быть измерены как информация... Таким образом, мы должны признать, что модель активационной теории эмоций 1967 г. строится на измерении неопределенности... Такую теорию лучше назвать «теорией неопределенности» или теорией зависимости эмоций от степени неопределенности» [Pribram, 1967, р. 833]. «Эмоции выражают отношения между восприятием и действием... Эмоции связаны с информационными процессами и механизмами контроля... На базе опыта эмоции возникают всякий раз, как только вероятность подкрепления действий представляется низкой» [Ibid., р. 836].
Заметим, что стадия прагматической неопределенности имеет место не только при формировании сложной системы условных реакций — динамического стереотипа, но и при формировании одиночного условного рефлекса, который также представляет систему, состоящую из двух или нескольких безусловных [Асратян, 1953, с. 208]. Еще в 1924 г. советский психиатр В.П. Осипов проницательно назвал первую стадию образования любого условного рефлекса — стадию генерализации — «эмоциональной» в отличие от более поздней «интеллектуальной, познавательной» стадии хорошо упроченного рефлекса [Осипов, 1924]. Эти две стадии (динамику ослабления, а затем и постепенного исчезновения эмоционального напряжения) легко продемонстрировать на примере выработки оборонительного условного рефлекса у человека. Испытуемых просили нажимать на ключ через 20 с после короткого звукового сигнала (рис. 1). Если субъект нажимал на ключ раньше 19 с после подачи сигнала или позднее 21 с, ему наносили на кожу предплечья болевое раздражение электрическим током порядка 60—90 В. После каждой пробы субъекта информировали о времени его реакции. Степень эмоционального напряжения измеряли по изменению частоты сердцебиений. Одновременно регистрировали кожногальванический рефлекс.
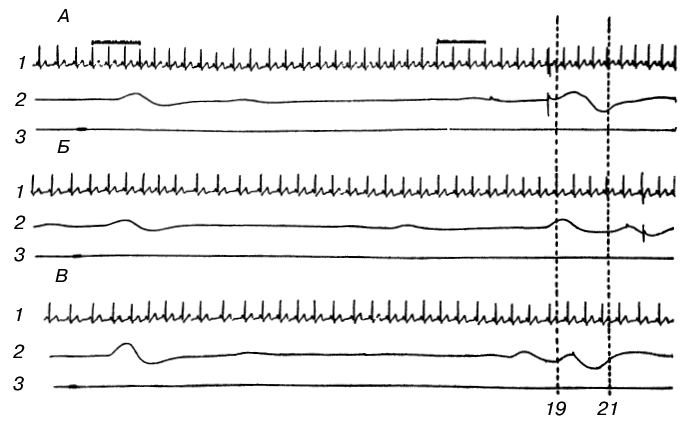
Рис. 1. Условный оборонительный рефлекс у человека
А, Б — ошибочные реакции, сопровождавшиеся болевым раздражением током; В — правильная реакция; 1 — ЭКГ; 2 — КГР, отметка реакции и тока; 3 — звуковой сигнал. Прерывистыми линиями отмечена «зона безопасности»
Нарастание частоты сердцебиений оценивали по суммарной продолжительности первых трех ударов сердца после звукового сигнала и последних трех ударов перед двигательной реакцией (скорость движения бумаги — 1,5 см/с). На протяжении первых 10 предъявлений условного сигнала субъект знал, что раздражения током не будет. Затем несколько раз применяли изолированное электрическое раздражение для того, чтобы определить интенсивность тока, не менее чем в три раза превышающую болевой порог. Эта интенсивность сохранялась постоянной на протяжении всего эксперимента.
На рис. 2 сплошная кривая показывает время двигательных реакций в последовательных пробах. Раздражения током наносились, когда ошибки субъекта превышали допустимое отклонение (обозначено двумя горизонтальными линиями). Прерывистая линия показывает изменения продолжительности шести сердечных сокращений в пересчете на число ударов в минуту. Верхняя горизонтальная линия обозначает среднюю частоту пульса в фоне до начала выработки условного рефлекса.
Если сравнить две стадии эксперимента, содержащие одинаковое число проб и одинаковое количество болевых раздражений током, можно убедиться, что суммарное отклонение частоты сердцебиений от исходного фона в этих двух частях эксперимента различно. Следовательно, оно зависит не только от количества наказаний. Опыты на 9 испытуемых показали, что суммарное изменение частоты сердцебиений пропорционально суммарному отклонению времени двигательных реакций от заданной величины, то есть пропорционально степени совершенства, точности и надежности условного оборонительного рефлекса (табл. 1). Это правило справедливо и для тех случаев, где величина ошибок (но не их количество, не количество болевых раздражений) нарастала вместе с нарастанием частоты сердцебиений и где, следовательно, динамику вегетативных сдвигов нельзя было объяснить привыканием к болевым стимулам по мере их повторения.
Эксперименты на животных также показывают, что мозг прогнозирует вероятность наказания в зависимости от степени совершенства инструментального двигательного рефлекса. Л.А. Преображенская вырабатывала условный оборонительный рефлекс у собак в ситуации,
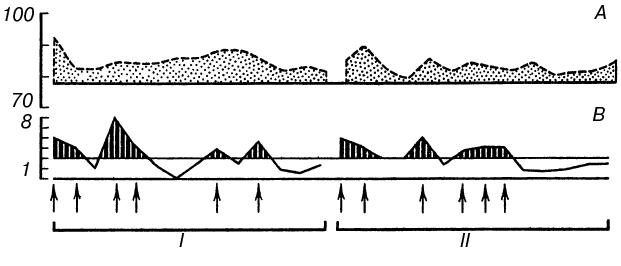
Рис. 2. Выработка условного оборонительного рефлекса
А — нарастание частоты сердцебиений по сравнению с фоном (уд/мин); В — отклонение времени двигательных реакций от заданной величины (с). Абсцисса — последовательные пробы на протяжении двух стадий эксперимента I и II. Стрелки — раздражения током
Таблица 1. Отношение суммарного изменения частоты сердцебиений к суммарной величине ошибок в опытах с девятью исследуемыми лицами
| Показатель |
Субъекты |
||||||||
| I |
II |
III |
IV |
V |
VI |
VII |
VIII |
IX |
|
| Число реакций в каждом из двух отрезков опыта |
8 |
14 |
9 |
9 |
15 |
8 |
17 |
9 |
10 |
| Число наказаний током |
2 |
6 |
3 |
3 |
8 |
5 |
7 |
7 |
3 |
| Отношение пульса и величины ошибок в первом отрезке |
7,5 |
1,56 |
3,28 |
5,4 |
3,08 |
0,48 |
0,66 |
0,42 |
0,73 |
| Отношение пульса и величины ошибок во втором отрезке |
6,1 |
1,55 |
3,11 |
5,8 |
3,19 |
0,46 |
0,83 |
0,40 |
0,61 |
где подъем передней лапы до определенного уровня и удержание ее на этом уровне в течение 10 с предотвращали болевое раздражение током противоположной задней лапы (методика Г.П. Зеленого, Г.В. Скипина, Р.Л. Винник и др.). Условный звуковой сигнал подавали за 10 с до болевого раздражения. Сочетание звука с болевым раздражением до выработки условного двигательного рефлекса вело к нарастанию амплитуды и процентного содержания тета-ритма в частотном спектре электрической активности дорзального гиппокампа (рис. 3). Количественный анализ обнаружил позитивную корреляцию изменений суммарного напряжения гиппокампального тета-ритма (измеренного по показаниям интегратора) с частотой сердцебиений. Оба симптома заметно ослабевали по мере стабилизации двигательного навыка, надежно избавляющего животное от боли. Любые затруднения в осуществлении движений вели к повторному нарастанию тета-ритма. Таким образом, опыты Л.А. Преображенской [1969; см. также: Konorski, Santibanez, Beck, 1968] показали, что интенсивность гиппокампального тета-ритма зависит не от двигательной активности самой по себе, но от эффективности двигательных актов, от их влияния на вероятность предотвращения болевых раздражений. Вместе с тем на степень эмоционального напряжения оказывает свое влияние и потребностно-мотивационный фактор: в опытах с оборонительными условными рефлексами суммарное напряжение тета-ритма и учащение сердцебиений были выражены значительно сильнее, чем в опытах с пищевыми.
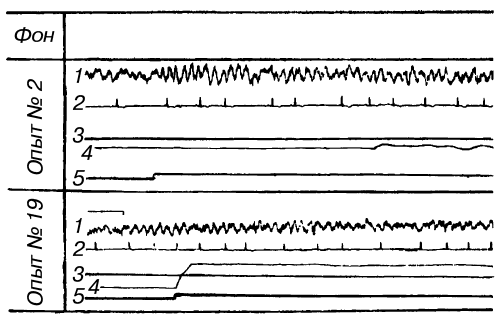
Рис. 3. Изменения гиппокампального тета-ритма и частоты сердцебиений во время выработки инструментального условного оборонительного рефлекса у собаки
1 — ЭГ дорзального гиппокампа; 2 — ЭКГ; 3 — уровень выключения тока; 4 — подъемы передней лапы, выключавшей ток; 5 — условный звуковой сигнал (по Л.А. Преображенской)
Заканчивая обзор данных, свидетельствующих об исчезновении эмоционального напряжения по мере выработки адекватной условной реакции, П. Фресс пишет: «Прежде всего следует подчеркнуть, что не существует эмоциогенной ситуации как таковой. Она зависит от отношения между мотивацией и возможностями субъекта» [Фресс, 1975, с. 133]. Предположение о том, что даже хорошо упроченный инструментальный оборонительный условный рефлекс продолжает мотивироваться страхом, как бы замещающим потребность избегания боли, подвергнуто аргументированной критике Ж. Нюттеном: «...потребность избегания болезненного стимула продолжает действовать (до тех пор, пока животное не усвоит, что сигнал не предвещает опасности), но поскольку животное не испытывает ни боли от стимула, ни угрозы опасной ситуации, оно не проявляет более эмоциональной реакции страха» [Нюттен, 1975, с. 70].
Литература переполнена экспериментальными данными, свидетельствующими о зависимости эмоционального напряжения от величины потребности (мотивации) и прогнозирования вероятности ее удовлетворения. Например, было установлено, что частота пульса у банковских служащих зависит от степени их ответственности (счет банкнотов различного достоинства) и количества информации, содержащейся в одной операции [Gantchev, Danev, Kitcheva, 1967]. Эмоциональные реакции обезьян закономерно зависят от изменения вероятности пищевого подкрепления [Melges, Popper, 1976]. По данным Д.Н. Меницкого и М.М. Хананашвили [1969], наибольшее эмоциональное напряжение у собак (визг, лай, чесание, царапанье кормушки) наблюдалось при вероятности подкрепления 1:4, а по мере продолжения опыта — при 1:2. Значение информационного фактора выступает особенно отчетливо в опытах со спаренными животными, когда оба партнера получают равное количество ударов током, но только один из них может предотвратить наказание соответствующей инструментальной реакцией. Показано, что именно у этого животного постепенно исчезают признаки страха, предотвращается изъявление слизистой желудка и кишечника [Desiderato, Newman, 1971; Jonas, Jonas, 1975; Starr, Mineka, 1977]. Мотивационный и информационный факторы генеза эмоционального напряжения имеют различный вес у разных животных одного и того же вида. По данным Л.А. Преображенской [1974], максимальное учащение сердечного ритма наблюдалось у одних собак при 30%-ном пищевом подкреплении, а у других — при 5%-ном. Следовательно, для собак второй группы суммарный «проигрыш» в удовлетворении пищевой потребности имел большее значение, чем неопределенность экспериментальной ситуации.
Имеются сведения и о том, что две составляющие эмоционального напряжения по-разному сказываются на величине различных вегетативных сдвигов. В опытах с участием человека величина побуждения (размер платы за правильное решение) преимущественно влияла на частоту пульса, дыхания и уровень электрического сопротивления кожи, а трудность задачи (количество выборов) — на объемный пульс и кожногальванический рефлекс [Wilkinson, El-Beheri, Gieseking, 1972]. Преимущественная связь кожногальванического рефлекса с информационным фактором обнаружена и в опытах, где кожногальванический рефлекс был слабее при ожидании болевого раздражения током с высокой вероятностью, чем при более редких, но трудно прогнозируемых ударах [Epstein, Bahm, 1971].
ОТРАЖАТЕЛЬНО-ОЦЕНОЧНАЯ ФУНКЦИЯ ЭМОЦИЙ
«Первые понятия, с которых начинается какая-нибудь наука, — писал Н.И. Лобачевский, — должны быть ясны и приведены к самому меньшему числу. Тогда только они могут служить прочным и достаточным основанием учения» [Лобачевский, 1976, с. 39]. Суммируя результаты собственных опытов и данные литературы, мы пришли в 1964 г. к выводу о том, что эмоция есть отражение мозгом человека и животных какой-либо актуальной потребности (ее качества и величины) и вероятности (возможности) ее удовлетворения, которую мозг оценивает на основе генетического и ранее приобретенного индивидуального опыта.
В самом общем виде правило возникновения эмоций можно представить в виде структурной формулы:
Э = f [П, (Ин – Ис), ...],
где Э — эмоция, ее степень, качество и знак; П — сила и качество актуальной потребности; (Ин – Ис) — оценка вероятности (возможности) удовлетворения потребности на основе врожденного и онтогенетического опыта; Ин — информация о средствах, прогностически необходимых для удовлетворения потребности; Ис — информация о средствах, которыми располагает субъект в данный момент.
Разумеется, эмоция зависит и от ряда других факторов, одни из которых нам хорошо известны, а о существовании других мы, возможно, еще и не подозреваем. К числу известных относятся:
� индивидуальные (типологические) особенности субъекта, прежде всего индивидуальные особенности его эмоциональности, мотивационной сферы, волевых качеств и т.п.;
• фактор времени, в зависимости от которого эмоциональная реакция приобретает характер стремительно развивающегося аффекта или настроения, сохраняющегося часами, днями и неделями;



















