

автордың кітабын онлайн тегін оқу Амшены, аргыны, атурая, ашкенази, канглы, их генетические родственники и соседи с древнейших времен до наших дней
Рахметолла Рахимжанович Байтасов
Амшены, аргыны, атурая, ашкенази, канглы, их генетические родственники и соседи с древнейших времен до наших дней
Шрифты предоставлены компанией «ПараТайп»
Благодарности:
Рахметолла Байтасов
© Рахметолла Рахимжанович Байтасов, 2022
В книге предпринята попытка найти истоки и проследить миграции носителей Y-хромосомной гаплогруппы G1. Для решения этой задачи использованы данные всех основных разделов археологической науки: первобытной археологии, восточной археологии, классической археологии, средневековой археологии, а также результаты новейших генетических исследований.
Полученные результаты будут интересны археологам, генетикам, историкам, любителям этнографии, генеалогии и истории.
ISBN 978-5-0056-1604-3
Создано в интеллектуальной издательской системе Ridero
Оглавление




ВВЕДЕНИЕ
Без знания истории своих семьи, рода, племени, народа невозможно укреплять духовность и ставить цели в жизни. Поэтому людям свойственно искать свои корни, истоки языка и культуры своей семьи и народа.
В настоящее время к семейными преданиям, архивным документам, добавился новый инструмент, позволяющий находить своих далеких предков — ДНК-генеалогия.
Я знал, что мои предки — казахские баи, репрессированные в 1930-е годы и подавляющее большинство родных и двоюродных братьев моего деда — «врага народа», как и сам дед, погибли на фронте в годы Второй мировой войны. Определив Y-хромосомную гаплогруппу узнал, что я потомок золотоордынского эмира Караходжи, жившего в конце XIV — начале XV вв. При этом, у меня очень редкая гаплогруппа — G1a. Поэтому появилась мысль найти истоки и проследить миграции носителей этой патрилинейной наследственности. Однако оказалось, что это очень трудная, почти невыполнимая задача, поскольку очень мало информации по палеоДНК и современным представителям данной гаплогруппы. Вместе с тем, опираясь на весьма скудные генетические данные и археологические материалы, удалось проследить маршруты миграции моих предков и их генетических родственников, а также ближайших соседей, начиная с эпохи палеолита вплоть до наших дней.
Результаты исследования излагаются в данном труде и будут интересны археологам, генетикам, историкам, любителям генеалогии, истории и этнографии.
Глава I ПРЕДЫСТОРИЯ СОВРЕМЕННОГО ЧЕЛОВЕЧЕСТВА
1.1. Homo bodoensis — непосредственный предок человека современного анатомического типа, Homo sapiens
Международная группа исследователей под руководством палеоантрополога Мирьяны Роксандич (Mirjana Roksandic) из Университета Виннипега (University of Winnipeg) описала Homo bodoensis, являющегося непосредственным предком современных людей. Вид назван по месту обнаружения черепа в 1976г., в долине реки Бодо Д'ар в Эфиопии. Череп принадлежал человеку, жившему в эпоху среднего плейстоцена, около полумиллиона лет назад.

Художественная реконструкция Homo bodoensis (Image credit: Ettore Mazza) [157]
Homo bodoensis обитал в Северной Африке и Юго-Восточной Европе.
В среднем плейстоцене[1], 774—129 тыс. лет назад[2], в Африке появился человек современного анатомического типа (Homo sapiens), а в Европе неандерталец (Homo neanderthalensis).
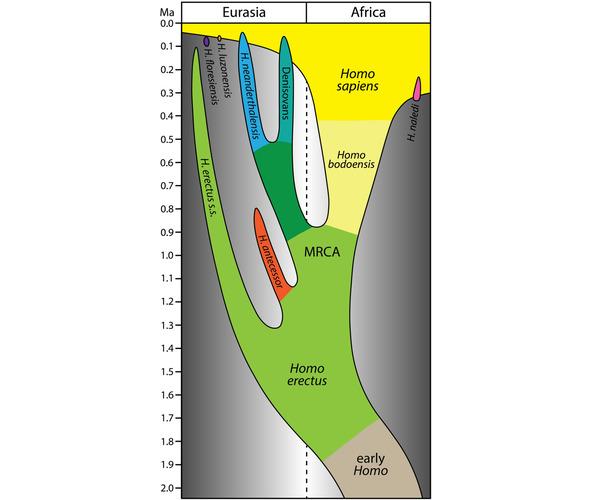
Упрощенная модель эволюции рода Homo в последние 2 млн. лет, в которой Homo bodoensis позиционируется как предок (в основном в Африке) Homo Sapiens [235].
По мнению палеонтологов, к Homo bodoensis следует относить почти все находки Homo, сделанные на африканском континенте и некоторые останки, обнаруженные на юго-востоке Европы.
Останки древних гоминид в Европе, классифицированные, как Homo heldelbergensis и rhodesiensis нужно считать неандертальцами [235].
В 2000-е годы большинство специалистов считало, что Homo sapiens возник в Восточной Африке примерно 200 тыс. лет назад, а современное внеафриканское человечество происходит от группы людей, которая покинула Африку через Баб-эль-Мандебский пролив примерно 65 тыс. лет назад. С территории Южной Аравии люди стали расселяться вдоль побережья Индийского океана на восток [16, c.21].
В 2012 году, генетики, исходя из расчётов скорости мутаций у современных людей, пришли к выводу, что время исхода из Африки следует удревнить до 130—90 тыс. лет назад [239].
Результаты исследования ДНК древних людей из Южной Африки, опубликованные в 2017 году, показали, что Homo sapiens появились на Земле ранее 300 тыс. лет назад [240].
Выход Homo sapiens из Африки, согласно Hershkovitz et al. (2018), мог произойти около 200 тыс. лет назад и ранее — останки человека современного анатомического типа из пещеры Мислия (Misliya Cave) в Леванте имеют возраст, полученный при совместном применении методов урановых рядов (Uranium-series, U-series) и электронно-парамагнитного резонанса (ЭПР)[3] 174±20 тыс. лет назад (календарных) (при стандартном отклонении 2σ) [186]. Однако, Sharp&Paces (2018) утверждают, что у Hershkovitz et al. (2018) нет достоверных U-series дат старше ~70 тыс. лет назад и останки имеют минимальный возраст не старше ~60—70 тыс. лет [244]. Отметим, что у российских археологов для пещеры Мислия приведена OSL дата[4] 130±33 тыс. лет назад (предположительно минимальная) [5, c.43].
1.2. Гибридизация человека современного анатомического типа (Homo sapiens) и неандертальцев (Homo neanderthalensis)
1.2.1. Неандертальцы
Неандертальцы (Homo neanderthalensis) — аборигены Европы, имеющие общего предка с человеком современного анатомического типа, жившего примерно 690—555 тыс. лет назад [91]. Согласно Я. В. Кузьмину (2020) возраст наиболее поздних неандертальцев Евразии составляет около 38,8—35,4 тыс. лет назад [80, с.132]. Использование методов ускорительной масс-спектрометрии[5] показало, что неандертальцы вымерли около 41030—39260 лет назад [187].

Реконструкция внешности неандертальского мальчика на основании черепа Гибралтар-2 (выполнена специалистами Цюрихского университета с использованием метода компьютерной палеоантропологии) [232]
Рисовали ли неандертальцы?
Несколько лет назад группа британских и немецких исследователей датировала уран-ториевым методом (²³⁴U/²³ºTh)[6] кальцитовые натёки на настенной живописи в трех испанских пещерах (Ла-Пасьега (Кантабрияна), Мальтравьесо (Экстремадура) и Ардалес (Андалусия)) и обнаружила, что её возраст — 64,8 тыс. лет. Поскольку в это время в Европе жили только неандертальцы, то по мнению учёных, рисунки оставлены ими [188].

Рисунки в пещере Ла-Пасьега (Испания), около 62 тыс. лет назад [216]
Однако археологи со всего мира усомнились в точности вышеприведенных дат, поскольку уран-ториевый метод мог дать ошибочное определение возраста кальцита[7]. В случае, если кальцитовые натёки не изолированы от окружающей среды, происходит выщелачивание урана и возраст, полученный уран-ториевым методом удревняется. Например, в пещере Ла-Пасьега для датировки рисунков из более чем 20 полученных дат, была выбрана самая древняя, сильно выбивающаяся из общего ряда и судя по всему, ошибочная из-за выщелачивания урана. Поэтому, по мнению исследователей, нет доказательств способности неандертальцев к рисованию [263].
Не подтверждают умение неандертальцев рисовать, исследования антропологов, которые обнаружили значительные расхождения в анатомии больших пальцев и кистей неандертальцев и современных людей.
Большой палец неандертальцев хорошо вытягивался и отклонялся в сторону, но плохо сгибался внутрь и двигался в стороны. Такая особенность кисти позволяла им крепко держать в руках орудия труда с длинными рукоятками (копьё, молот и т.д.), но затрудняла манипуляции с мелкими предметами, зажимая их между пальцами. Поэтому неандертальцы вряд ли могли вдевать нитки в иголки, хорошо шить [144], можно полагать и рисовать.
При этом, Кафзех 9 (Qafzeh 9), имеющий возраст 95 тыс. лет и считающийся ранним современным человеком, был способен, в отличие от неандертальцев, к более тонким и точным движениям пальцев [144].
Причины вымирания неандертальцев
Наиболее популярны три версии причин, вызвавших вымирание неандертальцев.
1) Сотрудники Института фундаментальных наук IBS (Южная Корея) с помощью суперкомпьютера построили модель, которая называет наиболее вероятной причиной вымирания неандертальцев — проигрыш в конкурентной борьбе за ресурсы с Homo sapiens. Возможными преимуществами современного человека перед неандертальцами называются использование более совершенных методов охоты, более сильный иммунитет, позволяющий противостоять новым заболеваниям и более высокий уровень рождаемости [254].
2) Одна из популярных гипотез — неандертальцы вымерли из-за того, что не смогли приспособиться к резкому изменению климата. Российские исследователи установили, что на Кавказе существует хронологический разрыв между концом среднего палеолита (ассоциируется с неандертальцами) и ранним верхним палеолитом (ассоциируется с Homo sapiens), который совпадает с событием Хейнрих 4, после которого в Кавказском регионе исчезают неандертальцы и появляются высокоразвитые каменные индустрии с преобладанием микропластинчатого компонента, сходные с ахмариеном Леванта [23, с. 171; 25; 26].
По мнению российских исследователей, мощные извержения вулканов, произошедшие практически одновременно на Апеннинах[8] и на Кавказе[9], привели в Северном полушарии к очень сильному похолоданию, по эффекту сопоставимому с ядерной зимой. Пепел покрывал огромные площади, его слои найдены в восточном Средиземноморье и на Кипре — более чем в 1500 км от извергавшегося вулкана. Следы извержений обнаруживаются в Восточной Европе от Пензы и Ростова на востоке до Южной Украины и Молдовы на западе. Возможно, «площадь поражения» была гораздо больше. Всё это привело к исчезновению неандертальцев [25; 118].
3) Исследование образцов, принадлежавших трём неандертальцам из пещеры Виндия в Хорватии, из Чагырской и Денисовой пещер на Алтае, а также одному денисовцу из Денисовой пещеры, показало, что причиной их вымирания могло стать низкое генетическое разнообразие и возможное распространение гемолитической болезни плода и новорождённого. В свою очередь, низкое разнообразие неандертальцев связано с их географической изоляцией, обусловленной климатическими изменениями в период плейстоцена. Эти факторы в сочетании с конкуренцией с Homo sapiens за пищевые ресурсы в одной и той же экологической нише, могли привести к исчезновению архаичных людей [159].
1.2.2. Время и регионы гибридизации современного человека и неандертальцев
В геномах, как неандертальцев, так и человека современного анатомического типа, обнаружены следы гибридизации друг с другом.
Человеческая ДНК в геномах неандертальцев: источники и время гибридизации
Генетические исследования свидетельствуют, что западноевропейский и кавказский неандертальцы (создатели индустрии восточный микок) имели общего предка около 150 тыс. лет назад [23, с.169].
Изучение ядерной ДНК неандертальцев и денисовцев показало, что они близкие родственники и их предок отделился от предка Homo sapiens 750—600 тыс. лет назад [176]. Установлено также, что неандертальцы и денисовцы скрещивались между собой, а денисовцы, видимо, скрещивались также, с Homo erectus, с которым они контактировали примерно 400 тыс. лет назад [199] и унаследовали от него 1% генома [192].
Сопоставление геномов двух неандертальцев (из Виндии и Денисовой пещеры), денисовца и двух африканцев, показало, что около 3% генома неандертальцев произошло от гибридизации между 200 и 300 тыс. лет назад с древней человеческой группой, ныне вымершей [192].
Сравнение митохондриальной ДНК неандертальца, жившего в пещере Холенштайн-Штадель (Hohlenstein-Stadel) на юго-западе современной Германии около 120 тыс. лет назад, 17 неандертальцев, живших около 430 тыс. лет назад в пещере Сима-де-лос-Уэсос (Sima de los Huesos) в Испании, трёх денисовцев и 54 современных людей, показало, что мтДНК образца из пещеры Холенштайн-Штадель сильно отличается от мтДНК более ранних неандертальцев и показывает сходство с человеческой. Генетики пришли к выводу, что неандертальцы приобрели человеческую митохондриальную ДНК примерно 220 тыс. лет назад, которая к 100 тыс. лет назад почти полностью заменила неандертальскую мтДНК. Исследователи предполагают, что популяция современных людей, вступившая в контакт с неандертальцами, впоследствии вымерла. МтДНК ранних неандертальцев сходна с таковой денисовцев [175; 176; 229].
Учёные также исследовали Y-хромосомы двух денисовских мужчин из Денисовой пещеры (Алтай, Россия), трёх неандертальцев из пещер Эль-Сидрон (Испания), Мезмайской (Кавказ, Россия) и Спи (Бельгия) и сравнили её с Y-хромосомой людей современного типа — африканцев и неафриканцев, представленных геномом человека из Усть-Ишима (Омская область), жившего 45 тыс. лет назад. Исследование показало, что в период от 350 до 150 тыс. лет назад произошла гибридизация неандертальцев с Homo sapiens, в результате чего неандертальцы получили от вымершей ветви человека современного анатомического типа Y-хромосому, которая заместила у них «родные» неандертальские варианты [226].
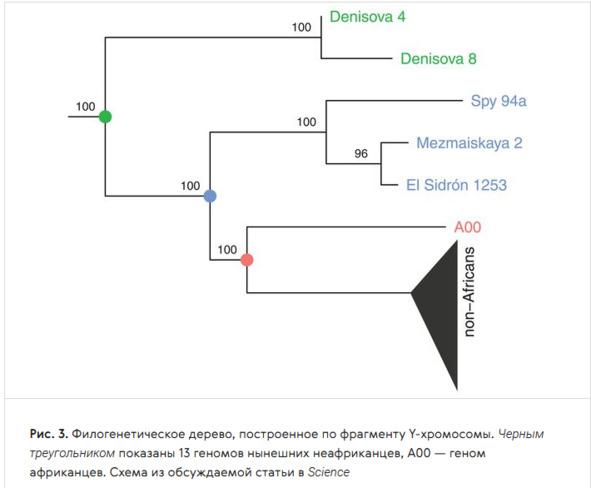
Филогенетическое дерево, построенное по фрагменту Y-хромосомы. Чёрный треугольник — 13 геномов современных неандертальцев; А00 — геном африканцев (Y-хромосомная гаплогруппа А00) [226]
Источники ДНК денисовцев у человека современного анатомического типа
В геноме жителей островов Меланезии (4—6% генома), а также в геномах популяций тихоокеанских островов, китайцев и японцев, есть следы гибридизации с денисовцами [160]. Больше всего денисовской примеси у папуасов. Эта примесь получена ими в результате гибридизации с дальними родственниками денисовцев, живших в Денисовой пещере на Алтае. Возможно, эта популяция денисовцев обитала на юге или юго-востоке Азии. Денисовская примесь есть также у жителей Южной Азии (индусы, бенгальцы и др.). Источник примеси у них тот же, что и у папуасов. Очень незначительная денисовская примесь есть у финнов [249]. Обнаружена денисовская примесь у исландцев, которая, по мнению исследователей, могла попасть к предкам исландцев от неандертальцев, до того, контактировавших с денисовцами либо далекие предки исландцев сталкивались с денисовцами [250].
У китайцев и японцев денисовская примесь получена из двух источников. Один из них, тот же, что и у папуасов, второй — денисовцы близкие к популяции, обитавшей в Денисовой пещере [154].
Время гибридизации современного человека и неандертальца
У европейцев приблизительно 2,5% [258], около 2,4% [207] генома имеет неандертальское происхождение.
M. Kuhlwilm et al. (2016) сравнив геномы неандертальцев, денисовского человека и современных людей, утверждают, что 125—120 тыс. лет назад группа людей современного анатомического типа вышла из Африки в направлении территории Среднего (Ближнего) Востока и Аравийского полуострова, где, по-видимому, обитали неандертальцы. В конечном итоге эта группа людей вымерла, но часть их генома унаследовали неандертальцы, которые мигрировали на Алтай. В геномах денисовца и двух неандертальцев (из Хорватии и Испании) следов ДНК древних африканцев нет, т.е. они не скрещивались с человеком современного анатомического типа.
60—50 тыс. лет назад другая группа людей современного анатомического типа вышла из Африки и встретилась с неандертальцами, мигрировавшими из Европы на Ближний (Средний) Восток и её представители спаривались с неандертальцами. Кроме того, потомки людей из этой группы скрещивались с денисовцами, ДНК которых обнаружена у меланезийцев и китайцев [199].
Chen et al. (2020), исследовав выборку из 2504 человек, происходящих из разных географических регионов, утверждают, что скрещивание современного человека с неандертальцами произошло один раз. Анализ показал, что у всех 504 исследуемых африканцев, неандертальское наследие совпадает с таковым у европейцев, а не у азиатов. Это соответствует сценарию миграции из Европы в Африку какой-то группы европейцев, после того, как их предки отделились от азиатов. Миграция, по-видимому, произошла в последние 20 тыс. лет. В целом, полученные результаты указывают на то, что скрещивание современного человека и неандертальцев происходило однократно, после выхода из Африки [155].
По мнению Browning et al. (2018), гибридизация современного человека с неандертальцами была один раз, либо повторная гибридизация, особенно в Азии, происходила с той же популяцией неандертальцев, с которой человечество столкнулось около 55 тыс. лет назад. Второй вариант более вероятен, так как у народов Азии достоверно больше неандертальской ДНК [154].
Vernot&Akey (2014) полагают, что неандертальцы и люди современного типа скрещивались в период между 75 и 40 тыс. лет назад [258].
С. В. Дробышевский (2019) исходит из данных по усть-ишимскому человеку, у которого неандертальская примесь составляет примерно 2,3%. Момент метисации оценивается в 13—7 тыс. лет до рождения усть-ишимского человека. Учитывая возраст самого усть-ишимца в 45 тыс. лет, смешение должно было происходить 58—52 тыс. лет назад [44]
Регион гибридизации неандертальцев и современного человека и представители Y-хромосомных гаплогрупп Homo sapiens, которые скрещивались с неандертальцами
Л. В. Голованова и В. Б. Дороничев (2010) доказывают, что в Европе неандертальцы и современный человек не скрещивались, поскольку неандертальцы вымерли в этой части света после 40 тыс. лет назад. Они отмечают, что самые поздние радиокарбоновые даты, указывают на существование неандертальцев в Европе незадолго до 38 тыс. лет назад — Фельдхофер в Германии, Эль Сидрон в Испании, Роше-де-Вильнев во Франции, Кюльна в Чехии, слой G3 в Виндии, Хорватия. Более поздние даты являются ошибочными (останки неандертальцев из слоя G1 в Виндии (29080±400 и 28020±360 радиокарбоновых лет) и слоя 3 в Мезмайской (29195±965 радиокарбоновых лет). В Мезмайской пещере возраст слоя 3 с находкой неандертальца определен на основании ЭПР-дат в интервале 73—63 тыс. лет назад. В пещере Виндия, «в слое G1, который формировался в течение длительного времени, вероятно, произошло смешение (палимпсест) стоянок неандертальцев и современных людей[10]» [24, с. 95].
Вместе с тем, результаты исследования палеоДНК древнейших европейцев, показывают наличие в их геномах ДНК неандертальцев и отсутствие связи с современным населением этой части света. Так, итоги исследования обитателей пещеры Бачо Киро (Болгария), живших 42,5—46 тыс. лет назад, у которых обнаружены следы гибридизации с неандертальцами за шесть-семь поколений до их рождения, свидетельствуют об отсутствии преемственности между создателями индустрии бачокирьена и современными европейцами. Гаплогруппы С1 (C-F3393) и F (F-M89), определенные у них, в настоящее время очень редки и встречаются с низкой частотой лишь в Юго-Восточной Азии и Японии [184].
Расшифровка генома современного человека, череп которого найден в пещере Златый кунь (Zlatý kůň) (Конепрусские пещеры, Чехия), показала, что это женщина, представительница митохондриальной гаплогруппы N. Жила она ~ 43 тыс. лет назад (~43 ka (95% highest posterior density = 31.5—52.6 ka)). Неандертальские гены предок этой женщины получил за 70—80 поколений до её рождения. Женщина из Zlatý kůň генетической преемственности с насельниками Европы, жившими после 40 тыс. лет назад, не имеет. Исследователи считают, что вероятной причиной этого может быть мегаизвержение Флегрейских полей и делают вывод, что большая часть неандертальских генов современным человеком получена в результате смешения с неандертальцами, родственными будущим европейским неандертальцам, а не обнаруженным на Горном Алтае [230].

Женщина из пещеры Златый кунь (Zlatý kůň) (Конепрусские пещеры, Чехия) [227]
S. Condemi et al. (2021), исследовав геномы неандертальцев, денисовца, папуасов, австралийцев-аборигенов и др., обнаружили, как они полагают, свидетельства гибридизации между левантийскими неандертальцами и Homo sapiens около 65 тыс. лет назад. По их мнению, гибридизация произошла во время раннего расселения Homo sapiens через Аравийский полуостров в направлении Австралии и Океании [159].
В исследовании M.A. Almarri et al. (2021) установлено, что в популяциях Ближнего Востока доля генетического вклада неандертальцев, в целом ниже, чем в популяциях Европы и Восточной Азии. По мнению исследователей, это обусловлено двумя причинами: неандертальские фрагменты размывались в ближневосточных геномах из-за смешения с африканской линией и из-за смешения с линией базальных евразийцев[11] (те и другие вовсе не имеют неандертальских генов). По этим двум причинам аравийские популяции имеют меньшую долю неандертальского вклада, чем левантийские [140].
Из результатов исследования M.A. Almarri et al. (2021) можно сделать ещё один вывод — гибридизации неандертальцев и современного человека на Ближнем Востоке не было. Косвенно данный вывод подтверждается тем, что в «пещере Кзал-Акир позднемустьерские и древнейшие верхнепалеолитические слои разделены седиментационной или эрозионной лакуной. Позднее мустье таких левантийских памятников, как Кебара или Тор-Фарадж, сильно отличается от древнейших комплексов верхнего палеолита» [82, с. 6].
Если исходить из того, что гибридизация неандертальцев и сапиенсов произошла около 55 тыс. лет назад [154], то с неандертальцами контактировали представители Y-хромосомных гаплогрупп С (сформировалась 65900 лет назад, время жизни последнего общего предка (TMRCA) 48800 лет назад) [267] и F (сформировалась 65900 лет назад, TMRCA 48800 лет назад) [267].
Гибридизация носителей гаплогруппы C с неандертальцами подтверждается тем, что представили её дочерних ветвей С1 и С2, имеют признаки смешения с этим древним человеком. В частности, нандертальская примесь обнаружена у носителей Y-хромосомной гаплогруппы С1 [248], живших в Сунгири (г. Владимир, Россия) 34654—33678 лет назад (калиброванная дата) [114], в Костёнках в Воронежской области России (геном человека Костёнки-14 (возраст 33250±500 некал. лет, 35—36 кал. тыс. лет [112, с.48]) содержит примерно 2,8% неандертальской примеси) [243] и др.
С гаплогруппой F не все так просто, поскольку существуют так называемые базальные евразийцы, у которых нет генов неандертальцев.
Jones E.R. et al. (2015) предполагают, что к базальным евразийцам (Basal Eurasian) относятся кавказские охотники-собиратели (Caucasus hunter-gatherer (CHG)) [194]. Их представители — люди из пещер Сацурблия (возраст 13300 лет; поздний верхний палеолит) и Котиас Клде (возраст 9895—9529 лет; эпоха мезолита). У мужчины из пещеры Сацурблия определены гаплогруппы — Y-хромосомная J1 [267] и митохондриальная К3; у мужчины из пещеры Котиас Клде (Kotias Klde cave) определены гаплогруппы — Y-хромосомная J2а и митохондриальная H13c. В настоящее время эти гапогруппы с высокой частотой встречаются в Грузии [194].
Вопрос о базальных евразийцах будет рассмотрен в отдельном разделе подробнее. Здесь же замечу, что базальными евразийцами, скорее всего, были носители Y-хромосомной гаплогруппы G. Тем не менее, поскольку Jones E.R. et al. (2015) считают обитателей пещер Сацурблия и Котиас Клде базальными евразийцами, можно предполагать, что у древних представителей гаплогруппы J также не было неандертальской примеси.
Определим, носители какой Y-хромосомной гаплогруппы контактировали с неандертальцами.
Гаплогруппа G — «внучка», а J — «прапраправнучка» гаплогруппы F. Материнской для J является гаплогруппа IJ, которая в свою очередь происходит из гаплогруппы IJK.
Поскольку IJ — материнская для гаплогруппы J, носители которой в древности не имели неандертальской примеси, то её представители, можно полагать, не заводили с неандертальцами «семейных» связей. Соответственно, можно предполагать, что с неандертальцами скрещивались представители Y-хромосомной гаплогруппы К (сформировалась 47200 (49700—44600) лет назад, TMRCA 45400 (49600—41400) лет назад), сестринской для IJ. При этом, носители гаплогруппы К должны были проживать в одном регионе с представителями гаплогруппы С (сформировалась 65900 (69100—62700) лет назад, TMRCA 48800 (51300—46400) лет назад) [267].
Данный вывод подтверждается тем, что неандертальская примесь (2,3%) есть у усть-ишимского человека (село Усть-Ишим Усть-Ишимского района Омской области), жившего около 45 тыс. лет назад [171], у которого определена базальная линия Y-хромосомной гаплогруппы K2a*-M2308 [228].
Неандертальская примесь обнаружена также у человека из пещеры Оасе (Пе́штера-ку-Оа́се (Peştera cu Oase), Румыния, более 40 тыс. лет назад), у которого определена, как и у усть-ишимского человека Y-хромосомная гаплогруппа К2а* [228] и митохондриальная гаплогруппа N [170]. Генетики полагают, что потомков Оасе 1 среди современных европейцев нет [228].

Оасе 1 — человек из пещеры Оасе (Пе́штера-ку-Оа́се (Peştera cu Oase), Румыния, более 40 тыс. лет назад) [227]. Первоначально, исходя из размера челюсти, исследователи предполагали, что Оасе 1 — неандерталец. Анализ ДНК его челюсти, найденной в 2002 г., показал, что у него до 11% неандертальских генов (у современных европейцев таких генов до 4%), что указывает на скрещивание с неандертальцами в предыдущих 4—6 поколениях [228].
У Prüfer K. et al. (2021) приведен возраст женщины из пещеры Златый кунь (Zlatý kůň) (Чехия) — ~43 тыс. лет назад. По мнению исследователей, неандертальские гены предок этой женщины получил за 70—80 поколений до её рождения [230]. Зная возраст женщины, проведем простейшие расчёты, учитывая при этом, что в древности женщины начинали рожать рано, в 12—13 лет. Мы возьмём для расчёта цифру в 15 лет. В этом случае получим: 80х15=1200 лет, 70х15=1050 лет. В итоге получим, что предок женщины, получивший гены от неандертальца, жил примерно 44 тыс. лет назад. В целом, не изменится картина, если в качестве возраста деторождения принять 20 лет: 80х20=1600 лет, 70х20=1400 лет. Время жизни предка женщины — получателя неандертальских генов составит около 45 тыс. лет.
Примечательно, что в этот период на Алтае появились карабомовская (43200±1500 лет назад (49797–44537 кал. лет назад) и 43300±1600 лет назад (49915–44614 кал. лет назад)) [107, с. 293]) и усть-каракольская (около 45 тыс. лет назад) [268]) верхнепалеолитические индустрии. Примерно такой же возраст имеют начальноверхнепалеолитические европейские индустрии богунисьен и бачокирьен, ближневосточный эмиран.
Отметим также, что гаплогруппа, к базальной линии которой относится усть-ишимец, является материнской для K-M2335 (TMRCA 41500 лет назад), от которой произошла гаплогруппа NO [267], предковая для N, преобладающей у уральских народов и О, доминирующей у китайцев и аустронезийских народов. У всех этих народов есть неандертальская примесь.
Поскольку M. Kuhlwilm et al. (2016) утверждают, что алтайские неандертальцы имеют гены современного человека и скрещивание произошло в районе Ближнего (Среднего) Востока 125—120 тыс. лет назад, обратим внимание на Алтай (заметим, что выше мы установили, что на Среднем Востоке (в Передней Азии) гибридизации неандертальцев и современного человека не было).
На Алтае стоянки неандертальцев найдены в трех пещерах: имени Окладникова, Страшной и Чагырской. Следы неандертальцев есть также в Денисовой пещере, где в эту же эпоху обитали денисовцы.
Палеогенетики из Института эволюционной антропологии Общества Макса Планка в Лейпциге (MPI-EVA) выделили ядерную ДНК неандертальцев не из костей, а из пещерного седимента (грунтовых отложений) в Денисовой и Чагырской пещерах на Алтае, а также в Галерии-де-лас-Эстатуас (Galería de las Estatuas (Галерея статуй)) в северной Испании. В этих пещерах, из слоёв возрастом 200—50 тыс. лет, были отобраны 150 образцов пещерного грунта. Для подтверждения верности определения использовали данные ядерной ДНК из костей и зубов и митохондриальной ДНК неандертальцев из тех же отложений (секвенированный геном женщины из Чагырской пещеры, полученный из фаланги пальца, и несколько образцов генома из Денисовой пещеры). В отложениях пещер Галерии-де-лас-Эстатуас и Чагырской обнаружена ДНК только неандертальцев, в Денисовой пещере — денисовцев и неандертальцев, без их смешения.
Геном неандертальцев из Чагырской пещеры существенно отличается от неандертальского генома из Денисовой пещеры. В испанской Галерии-де-лас-Эстатуас присутствуют следы неандертальцев, как из Чагырской, так и Денисовской пещер. По мнению исследователей, это связано с тем, что первая группа неандертальцев мигрировала из Европы в Азию 135 тыс. лет назад, а вторая — 105 тыс. лет назад [259].
Kolobova et al. (2020) установили, что ДНК неандертальцев, обитавших в Чагырской пещере 59—49 тыс. лет назад, свидетельствует об их более тесной связи с восточноевропейскими неандертальцами (Северная Хорватия и Северный Кавказ), чем с неандертальцами из пещеры Денисова, жившими 110 тыс. лет назад. Каменная индустрия из Чагырской пещеры тождественна инструментам микокской культуры эпохи среднего палеолита, распространенной в Восточной и Центральной Европе. Орудия из пещеры Окладникова (Сибирячихинской), расположенной восточнее Чагырской пещеры, имеющие радиоуглеродный возраст от 37,8 до 24,2 тыс. лет, также относятся к микокской культуре. В пещере Денисова, расположенной недалеко от Чагырской, артефактов микокской культуры нет.
Исследователи делают вывод, что неандертальцы приходили на Алтай дважды. Неандертальцы из Чагырской пещеры связаны со второй волной, которая пришла из Европы около 60 тыс. лет назад и принесла технокомплекс микок [195].
В другой работе этой же группы исследователей (Mafessoni F. et al. (2020)) утверждается, что неандертальцы из Чагырской пещеры пришли на Алтай из Европы около 80 тыс. лет назад и принесли специфическую микокскую традицию изготовления каменных орудий. Наиболее близкородственные чагырским неандертальцы обитали в Мезмайской пещере на Кавказе, в которой также нашли микокские каменные орудия. Установлено, что чагырская популяция имела менее 60 представителей и была изолированной [207].
Чагырские неандертальцы (микок) застали на Алтае денисовцев, которые ещё раньше контактировали с более древней волной неандертальцев, которые исчезли за 30 тыс. лет до прихода носителей микокской индустрии на Алтай.
Анализ ДНК девочки, считавшейся денисовкой, показал, что её отец — денисовец[12], а мать — неандерталка. Секвенирование генома чагырского неандертальца показало, что материнская линия у девочки представлена «чагырской» волной миграции, а в отцовской линии есть следы древних неандертальцев первой волны миграции.
Исследователи также утверждают, что все современные люди, кроме коренных африканцев, имеют в геноме около 2,4% генов, полученных в результате гибридизации с неандертальцами второй волны, к которой принадлежат хорватские (пещера Виндия) и чагырские неандертальцы. При этом, хорватские неандертальцы внесли больший вклад [207].
Исходя из вышеприведенной информации, можно сделать вывод, что неандертальские гены современные люди получили от неандертальцев с Балкан, создателей микокской индустрии. Однако, поскольку потомки древних людей (или их ближайших родственников) у которых обнаружена неандертальская ДНК, живут в настоящее время на востоке, в Азии, можно предположить, что гибридизация также происходила на территории промежуточной между Балканами и Алтаем. Миграцией с Балкан на восток древних европейцев, также можно объяснить больший процент неандертальской примеси у современных азиатских народов.
1.3. Создатели индустрии раннего и среднего левантийского мустье (Табун D, С) — предки внеафриканского человечества
Левантийское мустье по вариабельности, исходя из стратиграфической последовательности пещеры Табун, подразделяют на три фации — Табун D (раннее левантийское мустье), C (среднее левантийское мустье), B (позднее левантийское мустье.
Табун D
Hershkovitz et al (2018) отмечают, что для индустрии Табун D характерна развитая однополярная леваллуазская[13] технология, основанная на производстве удлиненных заготовок (пластин и леваллуазских острий). На ряде стоянок также использовались другие стратегии редукции нуклеусов. Например, на стоянках Рош-Эйн-Мор (Израиль) (около 200 тыс. лет назад), в пещере Мислия на склоне горы Кармель, в 12 километрах к югу от Хайфы (250—150 тыс. лет назад) применялась леваллуа-пластинчатая традиция. Каменные орудия Мислии, выполненные в технике Леваллуа, имеют сходство с орудиями такого типа с африканских стоянок Homo sapiens [186].
Отметим, что существует огромный разброс дат памятников Табун D. Так, К. К. Павленок (2015) пишет, что большинство комплексов мустье типа Табун D (стоянки Эль Коум, Умм-ель-Тлель, Рош Эйн Мор, Эйн Дифла, пещера Хайоним, Мислия и др.) относится к хронологическому интервалу 250—130 тыс. лет назад [95, с.188]. У А. П. Деревянко, М. В. Шунькова, C. В. Маркина (2014) приведена информация — ЭПР-даты индустрии типа Табун D находятся в пределах 230—120 тыс. лет назад, термолюминесцентные[14] — 280—230 тыс. лет назад [30, с. 44].
Hershkovitz et al. (2018) останки человека современного анатомического типа из пещеры Мислия (Misliya Cave) датируют возрастом 174±20 тыс. лет назад (календарных) (при стандартном отклонении 2σ), полученным при совместном применении методов урановых рядов (Uranium-series, U-series) и электронно-парамагнитного резонанса (ЭПР) [186]. Однако, Sharp&Paces (2018) утверждают, что у Hershkovitz et al. (2018) нет достоверных U-series дат старше ~ 70 тыс. лет назад и останки имеют минимальный возраст не старше ~ 60—70 тыс. лет [244].
Л. Б. Вишняцкий (2008) приводит следующие даты для памятников Табун D: пещера Мислия — минимальная ОСЛ дата 130±33 тыс. лет назад, стоянка Айн Дифла — ТЛ дата 105±15 тыс. лет назад и ЭПР даты от 102±13 до 162±18 тыс. лет назад, индустрия нижних слоев стоянки Бокер Тахтит — РУ даты (радиоуглеродные[15] даты) не более 50 тыс. лет [16, с. 60—61] и отмечает, что материалы «позднего левантийского мустье типа Табун С и В выглядят явно архаичней, чем гораздо более древние материалы мустье типа Табун D» [16, с. 61].
Дате раннего исхода человека современного анатомического типа из Африки не противоречат данные генетики. Так, согласно расчетам YFull, древнейшие в мире гаплогруппы A00 и A0-T, происходящие от «Y-хромосомного Адама», сформировались 235900 (243700—228300) лет назад. При этом, носители базальной линии гаплогруппы A00 проживают в настоящее время в Камеруне, время жизни их последнего общего предка (TMRCA) 37600 (42200—33100) лет назад [267]. Можно полагать, что носители клады A00 никогда Африку не покидали.
Сестринская к кладе A00, макрогаплогруппа A0-T, предковая для всего остального человечества мужского пола, сформировалась 235900 (243700—228300) лет назад, время жизни последнего общего предка (TMRCA) 161300 (169900—152900) лет назад [267]. Расчетный возраст макрогаплогруппы A0-T, почти совпадает с датировками раннего левантийского мустье (Табун D) по Hershkovitz et al. (2018) [186]. Поэтому исходя из возраста гаплогруппы и датировок Табун D можно предполагать, что индустрия Табун D оставлена Homo sapiens и, возможно, носителями гаплогруппы A0-T и её нисходящих линий, вплоть до ВТ (сформировалась 130700 (136400—125100) лет назад), TMRCA 88000 (92500—83600) лет назад) и СТ (сформировалась 88000 (92500—83600) лет назад, TMRCA 68500 (71800—65200) лет назад).
Y-хромосомная гаплогруппа ВТ (если определение выполнено верно), обнаружена у мужчины со стоянки Дольни-Вестонице (Dolní Věstonice; граветт Чехии, около 31 тыс. лет назад) [173]. В Дольни Вестонице определены также Y-хромосомные гаплогруппы СТ (notIJK), F [173] и C1a2 [210] (первоначально была определена как IJK [173]). Предварительно, как ВТ была определена гаплогруппа обитателя стоянки Буран-Кая (BuranKaya3A) в Крыму, жившего 36 тыс. лет назад, но дополнительные производные аллели указывали на то, что это может быть гаплогруппа СТ или С (не C1a2 или C1b). Обитатель Буран-Каи был носителем очень редкой митохондриальной гаплогруппы N1b, имеющей наибольшую концентрацию на Ближнем Востоке и распространенной на территории от Западной Евразии до Африки [150]. Данный факт указывает, что его предки родом с Ближнего Востока. Согласно Ancient DNA: https://ancientdna.info. Map based on public dataset on www.haplogroup.info. (www.indo-european.eu) у BuranKaya3A Y-хромосомная гаплогруппа C-CTS11043 (C1a) и митохондриальная гаплогруппа N1b и представляет он граветтскую культуру [143].
Гаплогруппа ВТ (ВТ-SRY10831.1) определена у двух хуэй (дунгане) [265]. Вполне может оказаться, что это одна из нисходящих линий ВТ (СТ и ниже).
По мнению A.E. Marks (1983), комплексы раннего левантийского мустье существовали вплоть до перехода к верхнему палеолиту — из мустье типа Табун D, продолжавшего развиваться в южных районах Леванта (пустыня Негев, Израиль) до 60—50 тыс. лет назад, происходит эмиран [208]. Л. Б. Вишняцкий полагает, что ввиду значительного хронологического разрыва с ранним левантийским мустье, индустрию стоянки Бокер Тахтит «следует рассматривать, по-видимому, не как продолжение последнего, а как результат вторичного и независимого появления в технологии и типологии переходных комплексов ряда тех же черт, что некогда были свойственны мустье типа Табун D» [16, с.60—61].
С ранним левантийским мустье типа Табун D ассоциируется появление платинчатой микроиндустрии [215] и предполагается связь населения Табун D с распространением леваллуа-пластинчатых индустрий в Евразии [101]. В частности, исследователи отмечают, что пластинчатая микроиндустрия характерна для слоя 23 палеолитической стоянки Кульбулак в отрогах Чаткальского хребта (Ташкентская область, Узбекистан), который предварительно датируется около 180 тыс. лет назад [101, с.68].
А. И. Кривошапкин (2012) считает, что из ближневосточного мустье типа Табун D происходит технология получения пластин с плоскостных и объемных нуклеусов, характерная для Оби-Рахмата (долина реки Пальтау, Бостанлыкский район Ташкентской области) в Узбекистане [70, с.27—28].
По мнению Е. П. Рыбина (2020), появление сходных типичных пластинчатых индустрий начального верхнего палеолита в Леванте и на Алтае является синхронным и, возможно, связано с «развитием на общей основе предковых леваллуа-пластинчатых индустрий Леванта типа раннелевантийского мустье представленных в комплексах Мислия и Рош-эйн-Мор» [107, с.498].
В. Ю. Коен и В. М. Степанчук (2001) прослеживают признаки технокомплекса Табун D, в кременецкой индустрии нижних слоев стоянки Кулычивка в Западной Волыни, датируемых около 31 тыс. лет назад [56, с.90—91]. При этом они отмечают, что нижний слой Кулычивки гетерогенен и представлен орудиями, типичными для технологически сходных индустрий (Богунисьен, Темната слой VI, Эмиран, а также с определенными оговорками леваллуа-мустье типа Табун D), а также ориньякским типологическим компонентом [56, с.91].
К раннему левантийскому мустье типа Табун D относятся, расположенные в горах Южной Осетии пещеры Кударо I, III и Цона (Южная Осетия); пещера Джручула в Имеретии (Западная Грузия); пещера Ховк I и стоянка открытого типа Баграташен I в Северной Армении [29, с.131]. По мнению Э. Ю. Демиденко (2020), возможная инфильтрация людей современного типа (раннее левантийское мустье типа Табун D) на Кавказ, «скорее всего, была эпизодическим и несистемным явлением, так как не привела к передвижению их по всему Северному Кавказу, не говоря уже про иные регионы Восточной Европы, где стоянок данных индустрий среднего палеолита нет» [29, с.133].
В. Б. Дороничев и Л. В. Голованова (2018) также отмечают близость к Табун D мустьерской индустрии кударо-джручульского типа на Южном Кавказе (пещеры Кударо 1, Кударо 3, Цона и Джручула) и исходя из того, что в пещере Джручула найден якобы зуб неандертальца[16], считают создателей кударо-джручульской индустрии неандертальцами. Кроме того, они говорят о сходстве с индустриями Южного Кавказа индустрий Северного Кавказа — в бассейне Терека (грот Сарадж-Чуко в Кабардино-Балкарии и пещера Мыштулагты Лагат (пещера Ласок) в Северной Осетии) [41, с.74].
Табун С
Табун С (Схул слой В, Кавзех слой XVII–XXIV, Доудара слой IIIB, Хайоним слой Е, Нар Ибрагим), фиксируется в Леванте в пределах 130–120 тыс. лет назад и существует вплоть до 75 тыс. лет назад [95, с. 188].
Для индустрии Табун С характерно преобладание широких овальных леваллуазских отщепов. С этим типом индустрии ассоциируются находки, как полагают некоторые исследователи, людей современного типа в Кафзех (110—90 тыс. лет назад и Схул[17] (130—100 тыс. лет назад) [181; 183].
Нужно отметить, что с периодом существования индустрии Табун С совпадает время формирования Y-хромосомной гаплогруппы ВТ — 130700 (136400—125100) лет назад и время жизни последнего общего предка (TMRCA) её носителей 88000 (92500—83600) лет назад [267].
В случае, если верны даты, приводимые Л. Б. Вишняцким (2008) для памятников Табун D: пещера Мислия — минимальная ОСЛ дата 130±33 тыс. лет назад, стоянка Айн Дифла — ТЛ дата 105±15 тыс. лет назад и ЭПР даты от 102±13 до 162±18 тыс. лет назад [16, с. 60—61], а не Hershkovitz I. et al (2018) — левантийского мустье типа Табун D, верхняя терраса пещеры Мислия — около 194—177 тыс. лет назад) [186], то гаплогруппа ВТ — патрилинейная наследственность представителей левантийского мустье D.
С. В. Дробышевский не считает группу Схул-Кафзех сапиенсами. По его мнению, они «не выходят за рамки видового размаха неандертальцев» и обозначает их P. neanderthalensis palestinensis [45].

Мужчина из погребения Схул-V. Графическая реконструкция Т. С. Балуевой [125] и изображение в цвете, выполненное по реконструкции художником Philip Edwin [227]
Отметим, что кисть руки Кафзех 9 (Qafzeh 9), имеющего возраст 95 тыс. лет и считающегося ранним современным человеком, отличалась от кисти неандертальцев, тем, что позволяла совершать тонкие и точные движения пальцами. Неандертальцы, в силу особенностей строения кисти, могли хорошо совершать действия с орудиями, имеющими длинные рукояти (копье и др.), но не могли манипулировать с мелкими предметами (вдевать нитки в иголки и т.п.) [144].
К. К. Павленок, Г. Д. Павленок, Р. Н. Курбанов (2020) констатируют, что доминирующей при изготовлении орудий в слое 23 стоянки Кульбулак (Узбекистан) была преференциальная техника леваллуа (preferentiаl Levallois method), распространенная в индустриях типа Табун С (стоянки Табун, Кафзех, Схул слой B, Хайоним слой Е и др.), относящихся к периоду от 190 до 85 тыс. лет назад. Преференциальная техника леваллуа соседствовала «с нуклеусами, расщепление которых велось в рамках параллельного пластинчатого метода, известного в регионе в первую очередь по материалам грота Оби-Рахмат» [101, с.68].
В слое 23 также представлена техника Нар-Ибрагим (Nahr Ibrahim technique), характерная для индустрий типа Табун С, результатом которой являются тронкировано-фасетированные изделия [101, с. 68].
Л. Б. Вишняцкий (2012) выдвинул предположение, что ранние сапиенсы Леванта (группа Схул-Кафзех, датируемая разными методами 81—115 тыс. лет до н.э.), которые, как считается, вымерли после 80 тыс. лет назад, могли мигрировать на юг Аравии, а затем в Австралию [19].
В пользу этой версии имеются археологические данные: комплекс Шибат Дихья I в районе Вади Сурдуд на Аравийском полуострове [162] и антропологические свидетельства. C. P. Groves (1994) указывал на определённое сходство черепа Кафзех 6 с черепами австралийских аборигенов, возможно сохранивших в длительной изоляции древний краниологический комплекс [182]. M. A. Schillaci (2008) проведя многомерный анализ морфометрических характеристик различных человеческих популяций пришёл к выводу, что палеолитическое население Австралии обнаруживает наибольшую генетическую близость к древнейшему населению Леванта. Кроме того, современное и верхнепалеолитическое население Европы имеет общего предка, отличающегося от древних жителей Австралии и древнейшего населения Леванта. Это, по его мнению, свидетельствует о том, что древнейшее население Леванта является непосредственным предком австралийцев-аборигенов или они имеют общего предка [242].
Несостоятельность гипотезы показывает то, что гаплогруппа C1-Z31885, наиболее часто встречающаяся у австралийцев-аборигенов сформировалась 47200 лет назад, время жизни последнего общего предка её представителей (TMRCA) 41100 лет назад. Вместе с тем, прародительская для C-Z31885 гаплогруппа С (сформировалась 65900 (69100—62700) лет назад, TMRCA 48800 (51300—46400) лет назад), является «внучкой» гаплогруппы СТ (сформировалась 88000 (92500—83600) лет назад, TMRCA 68500 (71800—65200) лет назад) и «дочерью» CF (сформировалась 68500 (71800—65200) лет назад, TMRCA 65900 (69100—62700) лет назад) [267]. Поэтому, можно предположить, что носители гаплогрупп СТ и CF внешне были похожи на австралийцев-аборигенов.
Дочерняя для СТ гаплогруппа CF и дочерняя для СF гаплогруппа F (сформировалась 65900 лет назад, TMRCA 48800 (51400—46300) лет назад) [267], судя по времени обитания ранних Homo sapiens в Леванте 250–130 тыс. лет назад [95, с. 188], 130–80 тыс. лет назад [16, с.60—61; 245], сформировались не в Леванте.
Табун В
В Леванте, в финале среднего палеолита 75/70—45 тыс. лет назад, во всех экологических зонах — средиземноморской прибрежной и аридной континентальной, господствовал леваллуазский технокомплекс [88, с. 13—14], который в этом регионе также называется мустье типа Табун В (стоянки Амуд (слой B1- 4), Кебара (слой VI—XII), Ксар Акил (слой XXVI), Тор Фараж (слой С), Тор Сабиха (слой С) и др.) [95, с. 188] и ассоциируется с неандертальцами [151; 223]. Эталонными технокомплексами мустье типа Табун В считаются ассамбляжи слоев X и IX пещеры Кебара. В основании слоя XI Кебары обнаружено погребение неандертальца [24, с.79].
Основой расщепления камня в левантийском мустье В являлась леваллуазская конвергентная техника [212; 231].
В. С. Славинский (2021) полагает, что среднепалеолитическая леваллуазская конвергентная однонаправленная техника для получения острий характерна для слоёв 19—21 грота Оби-Рахмат[18] в Узбекистане и оставлены эти слои неандертальцами [116, с.34]. Оби-Рахмат, по его мнению, был промежуточным пунктом на пути неандертальцев из Леванта на Алтай и далее на восток. Индустрии, содержащие артефакты леваллуазской конвергентной техники, фиксируются на территории Алтая в частности и, по всей видимости, Южной Сибири в интервале примерно 59—50 тыс. лет назад [116, с. 31—33].
Заметим, что происхождение алтайских неандертальцев, согласно недавно полученным данным, выводится из Европы [195; 207]. Все современные люди, кроме коренных африканцев, имеют в геноме около 2,4% генов, полученных в результате гибридизации с неандертальцами второй волны, к которой принадлежат хорватские (пещера Виндия) и чагырские неандертальцы. При этом, хорватские неандертальцы внесли больший вклад [207].
1.4. Влияние извержения супервулкана Тоба на популяции современного человека и неандертальцев
Вполне возможно, что неандертальцы появились на Ближнем Востоке, вследствие того, что многие районы на северо-западе Европы в это время стали малопригодными для обитания из-за извержения супервулкана Тоба на острове Суматра (Индонезия), произошедшего 77—69 тыс. лет назад [156; 217]. По результатам аргон-аргонового радиоизотопного датирования получена дата 75,0±0,9 тыс. лет [119].
Кроме Ближнего Востока, неандертальцы в этот период появились в Закавказье (пещеры Джручула и Сакажиа), в Передней Азии (пещера Шанидар), Средней Азии (пещеры Тешик-Таш, Оби-Рахмат, Ангилак), на Алтае (пещера Окладникова) и смежных с ним районах Центральной Азии [17].
Объём продуктов извержения супервулкана Тоба составил не менее 2,8 тыс. км³, из которых 800 км³ выпало в виде пепла. Пепел Тоба обнаружен в Южной Азии (в Малайзии мощность его слоя достигает 9 м, в Индии — 6 м,), в отложениях Южно-Китайского и Аравийского морей и Индийского океана и в Восточной Африке (микроскопическbq слой тефры вулкана Тоба обнаружен в донных отложениях озера Ньяса (Малави)). Следы извержения Тоба обнаружены в Гренландии, где в ледовых кернах, полученных в результате бурения ледового щита острова, фиксируется резкий скачок содержания солей серной кислоты, датированный 74.2±1.7 тыс. лет назад. Аналогичный скачок содержания серы зафиксирован во льдах Антарктиды.
С извержением Тоба связывают наступление вулканической зимы, — общего снижения температуры, вызванного экранирующим эффектом от выброшенных в стратосферу частиц пепла и сернистого газа, продлившейся не менее 6 лет, и очередного пика оледенения, продолжавшегося тысячу лет. Извержение супервулкана на Суматре могло привести к понижению температуры на 4—10° C в Европе, Центральной Азии и Северной Америке. На большей части территории Африки похолодание при этом не превысило 4° C.
Вероятно, в результате этого катаклизма вымерло население Южной и Юго-Восточной Азии. Преимущественные ветра не пустили пепел в Восточную Индонезию, а Гималаи и Тибет — в Центральную Азию, где выжили люди. В южных и северных областях Африки на стоянках, в период после извержения Тоба, есть перерыв в культурных отложениях, а число стоянок резко сокращается. Непрерывное накопление осадков наблюдается на некоторых центральноафриканских местонахождениях, свидетельствуя о том, что люди продолжали жить здесь постоянно [119; 141; 142; 193].
Согласно последнему исследованию истощения озонового слоя, вызванного выбросом сернистого газа и его преобразованием в сульфатный аэрозоль после извержения Тоба, установлено, что населявшие тропический пояс популяции, в том числе и ранние сапиенсы, могли серьезно пострадать не от похолодания, а вследствие экстремального ультрафиолетового облучения. Моделирование показало, что пиковый уровень истощения озонового слоя (в среднем на 20, а экваториальных широтах с 250 до 125 единиц Добсона) наступил через полгода после извержения Тоба. Через девять месяцев после извержения большая часть диоксида серы в облаке преобразовалась в аэрозоль, блокирующий поступление солнечного света, что понизило температуры в средних широтах, и без того охваченных похолоданием ледниковой эпохи, но не снизило температуру на экваторе. Однако недостаток озона в тропиках должен был привести к кратковременному, но экстремальному скачку УФ-индекса до значения 28[19].
Состояние озонового слоя над низкими широтами нормализовалось приблизительно через три года. Усиление канцерогенеза, подавление иммунной системы и общее повреждение ДНК должны были привести к значительной убыли численности человеческих популяций, а глазные заболевания и ожоги кожи в сочетании с нехваткой пищи — ещё более осложнить выживание [220].
Показательно, что время жизни последнего общего предка (TMRCA) мужчин, носителей гаплогруппы СТ, а это подавляющая часть современного человечества, — 68500 (71800—65200) лет назад (при том, что гаплогруппа сформировалась 88000 (92500—83600) лет назад) [267], совпадает со временем извержения супервулкана Тоба (77—69 тыс. лет назад [156; 217], 75,0±0,9 тыс. лет [119]) и появлением неандертальцев на Ближнем Востоке (75/70 тыс. лет назад [88, с. 13—14]).
Генетики отмечают, что в этот период человечество прошло «бутылочное горлышко (bottleneck)[20]» Так, согласно исследованию М. А. Almarri et al. (2021) в котором проанализированы 137 геномных последовательностей, относящихся к современным популяциям Леванта (сирийцы, иорданцы)), Ирака (иракские арабы, иракские курды) и Аравии (саудовцы, эмиратцы, йеменцы и оманцы), предковая ближневосточная популяция прошла через «бутылочное горлышко», 70—50 тыс. лет назад. Ко времени разделения на популяции Леванта и Аравии (20—15 тыс. лет назад) ее численность восстановилась [86; 140].
1.5. Атер, даббан, тарамсан, их происхождение и генофонд. Носители индустрии нубийского комплекса, как представители Y-хромосомной гаплогруппы DE. Прародина афразийцев
Согласно YFull YTree v9.04.00 [267] время формирования гаплогруппы ВТ 130700 (136400—125100) лет назад, время жизни последнего общего предка (TMRCA) 88000 (92500—83600) лет назад. Эти цифры очень хорошо совпадают со временем жизни ранних Homo sapiens в Леванте (130—80 тыс. лет назад) по J.J. Shea (2003) [245] и Л. Б. Вишняцкому (2008) [16, с.60—61].
Дочерними к ВТ являются линии В (сформировалась 88000 (92500—83600) лет назад, TMRCA 84800 (90300—79400) лет назад) и СТ (сформировалась 88000 (92500—83600) лет назад, TMRCA 68500 (71800—65200) лет назад). Линия В считается африканской, а СТ — внеафриканского человечества. Вместе с тем, время формирования дочерних для В гаплогрупп B-V2342 84800 (90300—79400) лет назад (TMRCA 26800 лет назад) и B-M8633 — 84800 (90300—79400) лет назад (TMRCA 83800 лет назад) [267], совпадает со временем исчезновения ранних Homo sapiens из Леванта. Это позволяет предположить, что гаплогруппа В появилась в Африке в результате ухода части древних сапиенсов из Леванта.
В Северной Африке в период перехода к верхнему палеолиту выделяются три культурные зоны — сахаро-атласская (атер), киренаикская (дабба или даббан) и нильская, — индустрии которых различались между собой по технико-типологическим характеристикам, имели разную историю и разные истоки [15, с.15].
Атер
Для атера типичны небольшие черешковые наконечники, датируемые около 90—60 тыс. лет назад, которые по мнению некоторых исследователей, являются наконечниками стрел (в Европе древнейшими вероятными наконечниками стрел являются наконечники стрелецкой культуры из Костёнок 1 и 4 с датировкой 35—24 тыс. лет назад) [47].
Около 80 тыс. лет назад у атерийцев появились символические предметы, свидетельствующие о поведении человека современного вида [30, с. 31]. В данном случае под символическими предметами понимаются раковины N. Gibbosulus с искусственными отверстиями, использовавшиеся как личные украшения. Они были обнаружены в атерийском горизонте пещеры Тафоральт (Grotte des Pigeons, Taforalt; 20 км от г. Беркан, Марокко), который имеет возраст, определенный на основании уранового, TL- и OSL- методов датирования, в интервале 91,5—74,4 тыс. лет назад (среднее значение 82,5 тыс. лет назад). До обнаружения раковин в пещере Тафаральт (Марокко) самыми древними такими находками со следами перфорации были раковины из пещер Бломбос (~78—75 тыс. лет; юг Африки, Капское побережье, юго-западнее Мосселбай), Сибуду (71 тыс. лет; пещера на севере провинции Квазулу-Натал в ЮАР), Схул и Кафзех (Израиль) [30, с.30]. В сентябре 2021г. опубликована статья, в которой утверждается, что 33 бусины из раковин, найденные в пещере Бизмун (юго-запад Марокко, южные склоны горы Джебель-Хадид) с другими артефактами атерийской культуры, имеют возраст, определенный методом урановых серий, 142—150 тыс. лет[21],. В то же время, атерийские слои датируются этим же методом ~62-≥142 тыс. лет [246].
Человеческие костные останки, обнаруженные в слоях с атером (Дар эс Солтан, Мугарет эль Алия, Зухра, Темара), принадлежат индивидам, явно отличавшимся от неандертальцев и приближавшимся по ряду важных краниофациальных характеристик к людям современного физического типа. В то же время, некоторым из них свойственны и отдельные неандерталоидные черты [15, с. 15].

Представитель атерийской культуры из Дар эс Солтана (Dar-es-Soltane). Реконструкция с сайта https://ancestralwhispers.org/reconstructions [161]
По мнению Л. Б. Вишняцкого (2006), корни атерийской культуры уходят в местное мустье [15, с.15]. «Некоторые исследователи связывают происхождение атера с индустрией люпембан Восточной и Центральной Африки» [31, с. 9].
Л. Б. Вишняцкий (2008) начало атера датировал 80—60 тыс. лет назад., а самые поздние памятники относил к периоду 30—22 тыс. лет назад [16, с.87].
А. П. Деревянко (2011) первоначально писал, что наиболее типичные и многочисленные атерийские местонахождения относятся к интервалу 40–20 тыс. лет назад (радиоуглеродные даты), но на северо-востоке Африки (в Киренаике) и в Египте атер датируется между 44700 и 43300 лет назад [33, с. 21]. Позже, А. П. Деревянко (2015) пишет, что применение новых методов датирования кардинально изменило ситуацию и удревнило атер. Видимо, формирование индустрии атер происходило около 112–110 тыс. лет назад, и существовала она длительное время. Так, для местонахождения Дар-эс-Солтан в окрестностях г. Рабат получена OSL-дата — 110 тыс. лет назад. Возраст памятников с аналогичной индустрией в районе г. Темара близок к этому значению. Образец из нижних атерских слоев в пещере Мугарет-эль-Алия датирован в пределах от 81±9 до 62±5 тыс. лет назад [31, с. 9].
Около 20 тыс. лет назад атер в Магрибе сменился финально-палеолитической иберо-маврской (оранской) культурой, «причём не исключено, что их разделяет несколько тысячелетий, когда не только Сахара, но и значительная часть средиземноморского побережья Северной Африки не имели постоянного населения» [16, с. 87].
На мой взгляд, каменная индустрия не может оставаться неизменной 90 тыс. лет (110 тыс. лет — 20 тыс. лет) и в датировках, удревняющих возраст атера столь кардинально могут быть ошибки. О том, что верны датировки начала атера, близкие 80 тыс. лет назад, свидетельствут появление у атерийцев около 80 тыс. лет назад символических предметов, что характерно для человека современного вида [30, с. 31]. Кроме того, изучив культурные и стратиграфические последовательности пещеры Рхафас на западе Марокко, некоторые ученые пришли к выводу, что переход от мустье к протоатеру в Северной Африке произошел 80—70 тыс. лет назад [30, с.30].
Можно предположить, что, возможно, население атерийской культуры было носителем гаплогруппы В (сформировалась 88000 (92500—83600) лет назад, TMRCA 84800 (90300—79400) лет назад) [267]. На это указывают даты бытования атерийской культуры, а также направление миграций атерийцев и наличие гаплогруппы В на Аравийском полуострове. В Саудовской Аравии есть базальная линия гаплогруппы B-M8498, время формирования которой 45200 (48300—42400) лет назад, TMRCA 45300 (48300—42400) лет назад [267]. Наиболее хорошо изученное местонахождение с атерийской индустрией на Аравийском полуострове находится на юго-западной окраине пустыни Руб аль Хали (в Северной Африке атер неизвестен к востоку от Нила). В Южной Аравии комплекс, содержащий орудия с черешком и бифасиальные формы, также известен в среднепалеолитической «индустрии Б» местонахождения Аль-Габр I в Западном Хадрамауте. «Наличие столь явных параллелей между каменными индустриями, существовавшими в расположенных на одной широте пустынях Северной Африки и Аравии, может объясняться только миграцией носителей атерских традиций в восточном направлении» [16, с. 82]. «Отсутствие каких-либо бы то ни было следов атера между Нилом и Красным морем заставляет предполагать, что миграция была разовой, скоротечной и, видимо, не очень масштабной…» [16, с. 80—82]. Радиоуглеродных дат по этим памятникам нет. По мнению исследователя памятника, он датируется от 35 до 20 тыс. лет назад [16, с.82].
Даббан
В Киренаике верхнепалеолитические комплексы культуры, именуемой дабба или даббан обнаружены в пещерах Хауа Фтеах и Хагфет-эд-Дабба. Cлой XX в пещере Хауа Фтеа датирован 31150 ± 40 лет назад, а слой XXVIII — 41000 лет назад. На основании этого Ч. Мак-Берни относил начало верхнего палеолита к 40–38 тыс. лет назад [30, с. 34]. Близкую древность имеет материал нижних слоев пещеры Хагфет-эд-Дабба, на что, помимо радиоуглеродной даты (38550±1600 BC), указывает и технико-типологическое сходство комплексов [14, с. 246].
По мнению археологов, смена мустье верхним палеолитом в Хауа Фтеах происходит внезапно [27, с.163; 257, p.109].
Даббан, по мнению Л. Б. Вишняцкого (2008), А. П. Деревянко и др. (2014), предположительно, можно рассматривать как вариант среднего или позднего ближневосточного эмирана [16, с. 91; 30, с. 34]. «Помимо общего и весьма оригинального „руководящего ископаемого“ (пластины с поперечной фаской), даббан сближает с этой индустрией и сохранение некоторых среднепалеолитических приемов в технологии (некраевое скалывание), и характер части скребков на отщепах» [16, с. 91].
Л. Б. Вишняцкий (2008) основную разницу между индустриями даббана и эмирана видит в том, что в Киренаике пластины с притупленным краем сосуществуют с пластинами с поперечной фаской (lames a chanfrein или pièces à chanfrein), а на Ближнем Востоке, в среднем эмиране пластин с притупленным краем практически нет и появляются они в позднем эмиране [16, с. 91]. Сопоставление даббана с индустрией нижних слоев Абу Халки[22] в Ливане, показало, что последняя несколько более архаична [16, с. 91].
Интересно, что пластины с поперечным резцовым сколом (lames à chanfrein), также называемые орудиями на пластинах с поперечной фаской (pièces à chanfrein), обнаруженные в даббане, и редко встречающиеся в палеолите, известны в Сибири как резцы диагонального типа и в Японии, как «резцы арайя»[23] [33, с. 22].
Антропологический тип носителей даббана неизвестен [15, с. 16].
О возможной миграции носителей культуры даббан из Азии в Африку свидетельствуют данные по исследованию мтДНК, которые показали, что у коренного населения некоторых северных и восточных районов Африки довольно часто встречаются два нетипичных для африканцев варианта мтДНК. Один из этих вариантов (M1) относится к группе M, которая распространена в Южной и Юго-Восточной Азии и в Океании. Второй вариант (U6) относится к распространенной по всей Евразии подгруппе R группы N. Генетики установили, что они происходят от евразийских групп M и N, но обособились от них около 40 тыс. лет назад. Более того, ближайшей родственницей гаплогруппы U6 является гаплогруппа U5, которая, по мнению многих экспертов, маркирует носителей культуры Ориньяк, проникших в Европу 45–40 тыс. лет назад [219].
Возникает вопрос: С какой Y-гаплогруппой пришли в Северную Африку M1 и U6? Поэтому рассмотрим генофонд современного населения Киренаики, данные по которому очень скудные.
У Ottoni C. et al. (2011) и Pereira L. et al. (2011) есть данные по туарегам, древнейшим жителям Киренаики. Наибольшие частоты у туарегов имеет гаплогруппа E1b1b, и, в частности, субклад E1b1b1b (E-M81) [222; 225]. У туарегов тахала в Ливии E1b1b составляет 100% генофонда [222].
Предполагается, что E-M81 возникла в Северной Африке около 14 тыс. лет назад [233] и одна её ветвь M183-PF2546 возрастом 2200 лет доминирует у северных и восточных берберов [267].
Кроме того, туареги, населяющие Нигер (5,6%) и Мали (9,1%), имеют в своем генофонде гаплогруппу Е-М78, носителями которой является население, разговаривающее на нигер-конголезских языках [225].
У туарегов тахала в Ливии [222] и туарегов Мали [225] есть гаплогруппа J1 (3%).
Вариант попадания гаплогруппы J1 в Африку с создателями левантийского эмирана исключается из-за молодого возраста гаплогруппы, которая сформировалась 31600 (34000—29200) лет назад, TMRCA 18300 (20300—16400) лет назад [267]. Время существования культуры Даббан — 31,5-38-40 тыс. лет назад (по оценкам разных авторов) [14, с. 246], эмиран имеет даты между ~46 тыс. лет назад и 36 тыс. лет назад [95, с. 187] и даже 50 тыс. лет назад [152].
У туарегов Аль Авайнат в Ливии обнаружены гаплогруппы R1b (8%) и G (3%) (остальной генофонд представлен гаплогруппой Е) [222]. Гаплогруппа G (1%) обнаружена также у туарегов Буркина-Фасо, у которых к тому же определена К-М9 (5,6%) [225].
Кроме того, у берберов Алжира обнаружена Y-ДНК гаплогруппа C1a2-V20 [266], характерная для населения ориньякской культуры, древнейшей в палеолите Европы. В настоящее время в Европе она почти вымерла.
Вполне возможно, что C1a2 попала в Африку из Испании 4,5 тыс. лет назад. Спорадические контакты между населением Пиренейского полуострова и Северной Африкой установлены ~ 2500 до н.э. [218]. Кроме того, C1a2, обнаруженная в Алжире, является нисходящей линией (образно говоря «внучкой») гаплогруппы C1-V20, представителями базальной ветви которой являются два брата, обозначенные как La Braña 1 и La Braña 2, жившие 7 тыс. лет назад в провинции Леон на северо-западе Испании [267]. Поэтому гаплогруппа C1a2 к даббану отношения не имеет.
Можно полагать, что гаплогруппа R1b в генофонд туарегов Аль Авайнат попала поздно, а гаплогруппы G и К-М9 (если она определена верно), представляют интерес в связи с даббаном и эмираном.
Что касается гаплогруппы G, то неизвестно, какая (ие) её клада обнаружена у туарегов. К тому же, существует огромный разрыв между (23,3 тыс. лет (48500—25200)) между временем формирования и временем жизни последнего общего предка носителей этой гаплогруппы. Интересно, что время существования культуры Даббан — 31,5—38/40 тыс. лет назад [14, с. 246], попадает в даты между временем возникновения гаплогруппы G и TMRCA. Если более глубокое исследование гаплогруппы G у туарегов покажет, что у них базальные или ещё неизвестные линии гаплогруппы G, то они могут оказаться потомками древних создателей индустрии даббан. Однако такое маловероятно.
Гаплогруппа К-М9 (сформировалась 47200 (49700—44600) лет назад, TMRCA 45400 (49600—41400) лет назад) происходит из гаплогруппы IJK. Её потомки — гаплогруппы LT, K2 [267], носителями нисходящих линий которых являются большинство современных мужчин Евразии и Америки.
Есть сомнения, что у туарегов определена именно К-М9. Вместе с тем, то, что резцы, подобные резцам даббана обнаружены на о. Хоккайдо, позволяет предположить, что население, использовавшее их, было родственным друг другу. Генофонд населения Северной Японии следующий (n=232): C1a1 — 1%, Ca2 — 4,5%, D1a — 2,5%, D1b — 61%, N — 1,5%, O1a — 0,5%, O2*/O2a — 2,5%, O2b — 15%, O3 — 12% [21]. C1a1 распространена на Окинаве (7%), Сикоку (10%) и Тохоку (10%), но, видимо, отсутствует на Хоккайдо и Кюсю [21]. К даббану она отношения не имеет, поскольку у берберов Алжира обнаружена C1a2, также не связанная с палеолитом Северной Африки.
Нас интересуют гаплогруппы дочерние к К-М9 (О, N, Q (есть в Восточной Японии (0,2% [21])).
Гаплогруппы О, N, Q — эт



















