

автордың кітабын онлайн тегін оқу Предыстория человечества: от «Адама» до «Авраама»
Рахметолла Байтасов
Предыстория человечества: от «Адама» до «Авраама»
Шрифты предоставлены компанией «ПараТайп»
© Рахметолла Байтасов, 2025
В книге на основе данных археологии, генетики, антропологии и лингвистики, рассматривается предыстория человечества от гипотетического «Адама» до гипотетического «Авраама» (Y-хромосомная гаплогруппа Е). Это делает книгу ценным источником для всех, кто интересуется происхождением человечества.
Автор не только излагает факты, но и анализирует противоречивые гипотезы, предлагая читателю взвешенные выводы.
Имеющиеся в книге рисунки и схемы помогают визуализировать сложные концепции.
ISBN 978-5-0067-0553-1
Создано в интеллектуальной издательской системе Ridero
Оглавление
Введение
Люди всего мира задаются вопросами: Кто мы? Кто наши предки и откуда они родом? Как долго существует человечество? По каким маршрутам передвигались наши предки до того, как оказались в современных местах обитания? Как установить происхождение отдельного человека, народа и их родство друг с другом? и т. д.
Пытаясь ответить на эти вопросы, многие люди уповают на потусторонние и сверхествественные силы, провозглашают человека венцом божественного творения, тем самым загоняя себя в тупик и признавая своё ничтожество (человек — тварь божья).
Есть категория людей, которые верят в человеческий разум, путём научного поиска стремятся познать окружающий мир, прошлое, настоящее и будущее человечества. К этой категории людей, которых можно назвать исследователями, относятся учёные, работающие в различных сферах науки. В настоящее время, науки, которые в наибольшей мере акцентированы на изучении предыстории человечества, — археология, генетика, лингвистика и др. В частности, археологи всего мира ищут признаки каменных индустрий, которые позволили бы различать среди них созданные людьми современного анатомического типа (ЛСА) (англ. Anatomically modern human, AMH, лат. Homo sapiens sapiens), неандертальцами и другими видами древних гоминидов. В последние годы достигнуты серьёзные успехи в этих поисках. Так, например, археологи видят различия в стратегиях первичного расщепления камня, применяемых нашими предками эпохи среднего палеолита, неандертальцами и денисовцами. В научной литературе всё более популярной становится идея о связи человека современного анатомического типа с индустриями, основанными на центростремительной технологии леваллуа и нубийском леваллуа.
В последние годы больших успехов достигли генетики. В частности, открытие новых методов изучения древней ДНК, позволило сравнивать геномы людей разных эпох, находить их генетических родственников в современных популяциях. Кроме того, исследование палеоДНК и современной ДНК позволяет верифицироватьь исторические реконструкции, основанные на археологических материалах.
В данной книге предпринята попытка, опираясь в основном на новейшие данные археологии и генетики заглянуть в предысторию человека современного анатомического типа от момента его формирования до появления и начала миграций представителей Y-хромосомной гаплогруппы Е. Автор не считает этот труд «истиной в последней инстанции». Где-то могут быть допущены ошибки, вызванные недостаточностью используемого материала, возможно неверным осмыслением некоторых данных, но, на мой взгляд, методология научного поиска по заданной тематике определена верно и послужит ещё многим исследователям в их благородной и трудной работе.
I. Непосредственный предшественник человека и его возраст
1.1. Возраст человека современного анатомического типа (Homo sapiens sapiens) и время его исхода из Африки
В 2000-е годы большинство специалистов считало, что Homo sapiens возник в Восточной Африке примерно 200 тыс. лет назад, а современное внеафриканское человечество происходит от группы людей, которая покинула Африку через Баб-эль-Мандебский пролив примерно 65 тыс. лет назад. С территории Южной Аравии люди стали расселяться вдоль побережья Индийского океана на восток [5, c.21].
В 2012 году, генетики, исходя из расчётов скорости мутаций у современных людей, пришли к выводу, что время исхода из Африки следует удревнить до 130—90 тыс. лет назад [37].
Результаты исследования ДНК древних людей из Южной Африки, опубликованные в 2017 году, показали, что Homo sapiens появились на Земле ранее 300 тыс. лет назад (самое раннее расхождение между человеческими популяциями могло произойти от 350 до 260 тыс. лет назад) [39]. Расчёты с помощью программного обеспечения BEAST2 показали, что время жизни последнего общего предка современных людей и неандертальцев (TMRCA) ~408 тыс. лет назад [20], т.е. мужчина, предок современного человечества, жил ~408 тыс. лет назад. Нужно отметить, что по мнению генетиков: «’«последний общий предок»», существовавший в точке генетического расхождения линий — это не конкретный индивидуум — отец или мать будущих «народов», «Адам» или «Ева», а условный момент в истории единой популяции, после которого свободный генетический обмен в ней прекращается» [6, с. 75].
Выход Homo sapiens из Африки, согласно Hershkovitz et al. (2018), мог произойти около 200 тыс. лет назад и ранее — останки человека современного анатомического типа из пещеры Мислия (Misliya Cave) в Леванте имеют возраст, полученный при совместном применении методов урановых рядов (Uranium-series, U-series) и электронно-парамагнитного резонанса (ЭПР) 174±20 тыс. лет назад (календарных) (при стандартном отклонении 2σ) [22]. Однако, Sharp, Paces (2018) утверждают, что у Hershkovitz et al. (2018) нет достоверных U-series дат старше ~70 тыс. лет назад и останки имеют минимальный возраст не старше ~60—70 тыс. лет [40]. Отметим, что у российских археологов для пещеры Мислия приведена OSL дата 130±33 тыс. лет назад (предположительно минимальная) [1, c.43].
1.2. Homo bodoensis или Homo heidelbergensis — непосредственный предок человека современного анатомического типа?
1.2.1. Гейдельбергский человек (Homo heidelbergensis)
Предком человека современного анатомического типа и неандертальцев (Homo neanderthalensis) долгое время считался гейдельбергский человек (Homo heidelbergensis) (рис. 1) (большинство исследователей придерживается этой точки зрения и поныне).

Рис. 1. Реконструкция головы взрослого Homo heidelbergensis. Выставлена в Зале происхождения человека Смитсоновского музея естественной истории в Вашингтоне, округ Колумбия (США) [23]
Название виду дано в честь г. Гейдельберг, вблизи которого (около деревни Мауэр), в земле Баден-Вюртемберг в Германии, впервые найдены останки этого гоминина. Время существования Homo neanderthalensis 800—30 тыс. лет назад. Рост Homo heidelbergensis, обнаруженных на юге Италии, судя по оставленным ими следам, не превышал 1,5 м [7].
В Европе, как полагает ряд исследователей, Homo heidelbergensis, стал предком неандертальцев, в Африке — современного человека. В Азии поздних Homo heidelbergensis (Шанидар, Табун, Тешик-Таш и др.) иногда относят к неандертальцам [2].
Wood, Collard (2000) cамым ранним представителем Homo heidelbergensis считают Бодо, который датируется примерно 600 тыс. лет назад (подробнее о Бодо, ниже). В этот вид древних людей они включают Флорисбад[1], Брокен Хилл (Кабве)[2] из Замбии, Эяси из Восточной Африки, Рабат[3] и Джебель-Ирхуд[4] из Северной Африки. Образцы среднего возраста Homo heidelbergensis (около 400 тыс. лет назад), по их мнению, включают черепа из Хоупфилда/Эландсфонтейна (человек Салданьа[5]) из Южной Африки, Ндуту из Танзании, Сале (Sale)[6] из Марокко и Thomas Quarry[7] из Марокко. Homo heidelbergensis в Азии, по их мнению, представлен свидетельствами из Нгандонга[8] в Индонезии, Дали, Мапа и Юньсянь (Yunxian) в Китае и Хатнора (Hathnora) в Индии [42, p.142].
Васильев (2009) сближает с Homo heidelbergensis (фактически включает в этот вид) гоминида из Бодо (Эфиопия) (550—640 тыс. лет). Имеющего эректоидно-сапиентные характеристики (большой надбровный валик, толстые кости свода, высокая чешуя височной кости, значительный изгиб основания черепа, намечающаяся клыковая ямка) Бодо относили то к архаичному Homo sapiens, то к Homo heidelbergensis. Также, как и Бодо, по мнению Васильева (2009) к Homo heidelbergensis относятся гоминиды из Восточной и Южной Африки — Ндуту (370—990 тыс. лет назад), Брокен Хилл (130—300 тыс. лет), Салданья (400—700 тыс. лет), Эяси (130 тыс. лет) [4, с. 84]. К Homo heidelbergensis Васильев (2009) относит также среднеплейстоценовых поздних гоминид Африки, которых некоторые авторы выделяют в вид Homo helmei — Элие-Спрингс[9], Летоли 18 (около 200 тыс. лет), Омо 2, Флорисбад (260 тыс. лет), Бордер Кэйв, Мумбва и др. [4, с. 84].
В Леванте и Передней Азии Васильев (2009) относит к Homo heidelbergensis Кафзех VI и IX (115—92 тыс. лет), Схул IV и V (100—70 тыс. лет), Табун I (120 тыс. лет), Амуд I (50—40 тыс. лет), Шанидар I и II (46 тыс. лет), Кебара (61—48 тыс. лет). Обосновывает он такое мнение морфлогической мозаичностью этих людей (сочетание эректоидности с сапиентностью и/или неандерталоидностью) [4, с.85]. По мнению Васильева (2009), поскольку Передняя Азия представляет территорию, по которой постоянно проходили миграции древних людей из Африки на восток и с востока в Европу, здесь изначально Homo ergaster, а позднее Homo heidelbergensis, имея в своей морфологии некоторые сапиентные и/или неандерталоидные характеристики, формировали различные метисные варианты, максимально стабилизирующиеся только на подвидовом уровне. Исходя из этих соображений Васильев (2009) относит неандерталоидно-сапиентного Схула, эректоидно-неандерталоидных Табун, Шанидар, Амуд и эректоидно-сапиентного Кафзех VI к Homo heidelbergensis, определяя их таксономический статус по виду предшественнику, полагая, что практически во всех случаях, предшественником переднеазиатского полиморфного населения был вид Homo heidelbergensis (один из представителей, наряду с Homo erectus и Homo ergaster, эректоидных форм) [4, с. 85].
К Homo heidelbergensis могут относиться гоминиды из Китая — Дали (120—230 тыс. лет) и Цзиньнюшань (200—300 тыс. лет), а также два черепа, найденные в округе Юнсян (провинция Хубэй). Кроме того, к Homo heidelbergensis относятся люди из Нгандонга (примерно 100 тыс. лет). [4, с. 85].
Зубов (2001) описывает эволюцию гейдельбергского человека в Европе в неандертальцев и в современного человека в Африке следующим образом: «Одна из ветвей этого вида примерно 800 тыс. лет назад мигрировала в Европу и, адаптировавшись к довольно суровому климату новых мест, постепенно преобразовалась в Homo neanderthalensis, другая ветвь осталась в Африке и дала начало эволюционной линии Хоупфилд — Ндуту[10] — Кабве — Элие Спрингс- Омо-2[11] — LH18 (Летоли-18[12] или Нгалоба; ~120 тыс. лет назад) из Танзании) и, наконец, Омо-1 (Homo sapiens)[13]. Впоследствии потомки этой линии мигрировали в Переднюю Азию, а затем — примерно 35—40 тыс. лет назад — в Европу» [8, с.].
1.2.2. Homo bodoensis
В 2022 году международная группа исследователей под руководством палеоантрополога Мирьяны Роксандич (Mirjana Roksandic) описала Homo bodoensis (рис. 2), который, по их мнению, является непосредственным предком современных людей. Вид назван по месту обнаружения черепа в 1976 г., в долине реки Бодо Д'ар в Эфиопии. Череп принадлежал человеку, жившему в эпоху среднего плейстоцена, около полумиллиона лет назад [35]. Выше показано, что этого человека также относят к самым ранним представителям вида Homo heidelbergensis [42, p.142].

Рис. 2. Художественная реконструкция Homo bodoensis (Image credit: Ettore Mazza) [16]
Homo bodoensis обитал в Северной Африке и Юго-Восточной Европе. В среднем плейстоцене, 774—129 тыс. лет назад, в Африке появился человек современного анатомического типа (Homo sapiens), а в Европе неандерталец (Homo neanderthalensis) (рис. 3) [35].
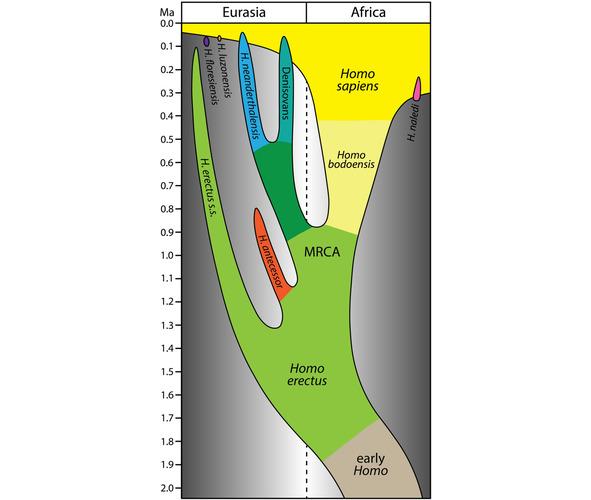
Рис. 3. Упрощенная модель эволюции рода Homo в последние 2 млн. лет, в которой Homo bodoensis позиционируется как предок (в основном в Африке) Homo Sapiens [35, Fig. 1].
По мнению Roksandic et al. (2022), к Homo bodoensis следует относить почти все находки Homo, сделанные на африканском континенте и некоторые останки, обнаруженные на юго-востоке Европы. Останки древних гоминид в Европе, классифицированные, как Homo heldelbergensis и rhodesiensis нужно считать неандертальцами [35].
Вполне возможно, что творением рук Homo bodoensis являются деревянные постройки в районе водопада Каламбо Фоллс (Kalambo Falls) в Замбии (граница Замбии и Танзании). Археологи, в заболоченных отложениях на месте археологических раскопок близ этого водопада (местонахождение Каламбо Фоллс), обнаружили два переплетающихся бревна из ивы крупноплодной (Combretum zeyheri), соединённых поперек намеренно вырезанной выемкой (обнаружены следы обработки огнём и каменными инструментами). Исследователи предполагают, что брёвна могли быть частью фундамента сооружения, дорожкой или приподнятой платформой на участке, который регулярно заливался водой.
Датирование этих брёвен методом люминесцентного датирования дало дату не менее 476±23 тыс. лет назад. Эта конструкция не имеет известных параллелей ни в африканском, ни в евразийском палеолите. Самым ранним известным деревянным артефактом является фрагмент полированной доски из ашельской стоянки Гешер Бенот Яаков, Израиль, возрастом более 780 тыс. лет назад. Деревянные орудия для собирательства и охоты появились около 400 тыс. лет назад в Европе, Китае и, возможно, в Африке. В Каламбо также обнаружили четыре деревянных орудия возрастом от 390 до 324 тыс. лет назад, в том числе клин, палку-копалку, обрезанное бревно и надрезанную ветку [13].
Человеческие останки на месте находки деревянных артефактов обнаружены не были, но исследователи предполагают, что они были созданы Homo heidelbergensis, обитавшим в этот период в данном регионе Африки [14; 17].
Исходя из возраста брёвен в Каламбо Фоллс 476 ± 23 тыс. назад, можно согласиться с тем, что их обработкой занимались Homo heidelbergensis или, если придерживаться гипотезы Roksandic et al. (2022), что на мой взгляд, более верно, Homo bodoensis, который, по их мнению, является непосредственным предком современных людей [35]. Выше упоминалось, что расчёты с помощью программного обеспечение BEAST2 показали, что время жизни последнего общего предка современных людей и неандертальцев (TMRCA) ~408 тыс. лет назад [20]. Если исходить из даты ~408 тыс. лет назад, то обитатели Каламбо Фоллс — общие предки человека современного анатомического типа и неандертальцев. В то же время, деревянные орудия, возрастом от 390 до 324 тыс. лет назад в Каламбо [13], — творение предков человека современного типа. Они созданы после ухода предков неандертальцев в Европу и, возможно, на север Африки (Джебель Ирхуд).
Брокен-Хилл, Кабве / Broken Hill, Kabwe (Родезийский человек; Rhodesian Man) — череп гейдельбергского человека (Homo heidelbergensis), найденный в Юго-Западной Замбии (в прошлом — Северная Родезия) (г. Кабве, близ станции Брокен-Хилл, свинцово-цинковый рудник Брокен-Хилл). По данным датирования методом урановых рядов имеет возраст 299±25 тыс. лет [19].
Флорисбад — местонахождение под открытым небом с артефактами среднего каменного века (MSA) в провинции Фри-Стейт (Free State) Южная Африка (в 45 км к северо-западу от Блумфонтейна (Bloemfontein), озеро Хагенштедт). Культурные комплексы включают ранний MSA, датированный 279±47 тыс. лет назад, сильно ретушированную форму MSA возрастом 157±21 тыс. лет назад и минимально ретушированный комплекс MSA из ряда заселённых горизонтов возрастом 121±6 тыс. лет назад. Здесь обнаружен череп раннего Homo sapiens, датированный 259±35 тыс. лет назад (электрон-спин-резонанс на человеческом зубе, ассоциированном с черепом) [25; 26]. Scerri et al. (2018) рассматривают Флорисбад как представителя одной из линий ранних Homo sapiens, сформировавших в результате гибридизации и эволюции человека современного анатомического типа [38].
Согласно результатам последних исследований Ирхуд 1 — Homo heidelbergensis [3, с.105], а археологические слои в Джебель Ирхуде датируются от 240±35 до 378±30 тыс. лет назад, в среднем 315±34 тыс. лет назад (термолюминесцентные даты). Непосредственно на останках ребёнка Ирхуд 3 получена дата 286±32 тыс. лет назад [24].
Рабат — верхняя челюсть с зубами. Обнаружена в Марокко, южнее г. Рабата, в карьере Кебибат, в литологическом слое S2, для которого получена дата 137±7 тыс. лет назад. Исследователи полагают, что этот слой был сформирован между концом MIS 7 и, вероятно, концом MIS 6. Останки человеческого черепа, найденные в Кебибате, принадлежат прибрежной дюне, образовавшейся во время MIS 6. С палеоантропологической и геохронологической точки зрения останки человека из Кебибата характеризуются более архаичной морфологией и являются более древними, чем другие останки человека, найденные в прибрежных пещерах Темары). Более того, сравнение внешнего вида черепов человека со стоянки Кебибат и стоянки Джебель-Ирхуд позволяет предположить, что морфологически в стоянке Джебель-Ирхуд черепа менее архаичны и относительно более древние, их возраст составляет 300 тыс. лет [15].
Сале, Эль Хамра (El Hamra) — предполагаемый возраст 400 тыс. лет назад [36].
Хоупфилд (Салданья (Saldanha, Hopefield 1, Elandsfontein)) — черепная коробка мужчины найдена в Эландсфонтейне, Салданья (Южная Африка). Предполагаемый возраст: 150–400 тыс. лет (?). Точная датировка слоёв местонахождения Салданья невозможна. Сопутствующие орудия относятся к нескольким культурным традициям: финальному ашелю, среднему каменному веку и позднему каменному веку. Таксономическое определение: Homo erectus soloensis (нгандонгский, или солойский человеr) или Homo heidelbergensis. Исследователи отмечают похожесть этого черепа на Брокен-Хилл 1 (Кабве) [11].
Нгандонг, местонахождение ископаемого человека (явантроп; от 550 до 25 тыс. лет назад) на о. Ява (Индонезия). На левом берегу р. Соло (=Кали-Бенгаван) близ села Нгандонг обнаружены черепные крышки и берцовые кости 12 человек: 8 взрослых (мужчины: Соло 5, 9, 10, 11; женщины: Соло 1, 4, 6, 8), 1 ребёнка мужского пола (Соло 2) и 3 — неопределённого возраста. Почти у всех черепов было проломлено основание (вероятно, признак каннибализма). Индивид Нгандонг V (он же Соло 4) и три других были убиты ударом по голове. Найдены также грубые каменные отщепы, пластины, каменный шар; костяные нож с отполированным краем и гарпун, остроконечное орудие из оленьего рога. Возможно, орудия и фауна с людьми из разных слоёв. Мозаика эректоидных и сапиентных черт, сходство с черепом из Кабве позволяют отнести его к гейдельбергскому человеку, а поздний возраст заставляет считать его реликтовой популяцией этого вида. Некоторые зарубежные исследователи рассматривают гоминид из Нгандонга как предков современных австралийских аборигенов [9]. Hawks et al. (2000), провели поиск потенциального предка WLH-50 среди трёх популяций (Нгандонг, африканцы позднего плейстоцена, гоминиды Леванта из Схула и Кафзеха). В результате они пришли к выводу, что гоминиды Нгандонг или подобная им популяция, наряду с двумя другими группами, внесли значительный вклад в родословную WLH-50 и современных австралийцев-аборигенов [21].
Обнаруженные в пещере Thomas Quarry I окаменелости гомининов, явно близки к предковой линии Homo sapiens [33].
KNM-ES 11693 или Eliye Springs (Элие-Спрингс) — череп взрослого человека, найденный на северо-западе Кении (деревня Элие-Спрингс на западном берегу озера Туркана). Эта находка не имеет чёткого стратиграфического положения и датируется ориентировочно 300–200 тыс. лет назад. На сайте Смитсоновского Национального музея естественной истории (Вашингтон, США) определён как Homo heidelbergensis [18]. Некоторые исследователи относят его в промежуточный между гейдельбергским человеком и Homo sapiens вид Homo helmei [12].
Часть верхней челюсти и практически неповреждённый свод черепа человека Нгалоба (Ngaloba) LH 18 найден на местонахождении Нгалоба (Ngaloba) в местности Лаэтоли (Laetoli), вблизи озера Эяси (Lake Eyasi), в 50 км южнее Олдувая, в Танзании. Слои Нгалоба в которых он был обнаружен датируются примерно 120±30 тыс. лет назад [28]. На сайте Смитсоновского Национального музея естественной истории (Вашингтон, США) он отнесён к виду Homo sapiens, т.е. к человеку современного анатомического типа и датирован — около 120 тыс. лет [32]. Magori, Day (1983), изучавшие череп Ngaloba LH18 отметили наличие на нём смеси как архаичных, так и современных особенностей (например, толстые надглазничные торы (supraorbital tori), которые тем не менее чётко разделены на медиальный и латеральный отделы, общая ширина черепа и изогнутая затылочная часть). Объём черепа около 1200 см³. В морфологии LH 18 есть некоторые различия от черепов других ранних африканских Homo sapiens, но достаточно сходств, свидетельствующих о принадлежности их людям одного вида. LH 18 совершенно не похож на изученные черепа человека прямоходящего и неандертальца. Метрический анализ подтверждает связи между LH 18 и другими архаичными черепами Homo sapiens из Африки, особенно Эяси и Омо 1 [28]. По мнению Lahr et al. (2016) присутствие на черепе LH 18 как архаичных, так и современных особенностей может указывать либо на большую морфологическую изменчивость раннего человеческого населения Восточной Африки или на существование нескольких типичных групп в регионе в то время [27]. В исследовании Mounier, Lahr (2019) восточноафриканский LH18 продемонстрировал наименьшее сходство с современными людьми. Североафриканский Ирхуд-1 занял промежуточное положение между современными людьми и неандертальцами, а южноафриканский Флорисбад и, в меньшей степени, восточноафриканские KNM-ES 11693 и Omo II, показали наибольшую близость к современным людям [31].
Омо 1 из формации Кибиш в Эфиопии, возрастом 232±22 тыс. лет назад, считается древнейшим известным человеком современного анатомического типа [41].
Ндуту 1 (Ndutu 1) — череп, найденный вблизи озера Ндуту (Танзания, юго-восточная часть плато Серенгети, близ вулканического кратера Нгоронгоро, около западного конца Олдувайского ущелья). Форма реконструированного свода черепа Ндуту оказалась наиболее близкой к Sima de los Huesos (SH) 5, в то время как его сагиттальный профиль был наиболее похож на профиль Кабве. Предполагаемый возраст — около 450 тыс. лет назад [30].
Омо 2 (Omo II) — черепная коробка, найденная в формации Кибиш, в Эфиопии (Kibish formation, Ethiopia), по общей форме напоминающая черепа из Нгандонга. Предполагаемый возраст: 104-195-196 тыс. лет назад (⁴⁰Ar/³ꝰAr; 2008 год) [29].
Флорисбад — местонахождение под открытым небом с артефактами среднего каменного века (MSA) в провинции Фри-Стейт (Free State) Южная Африка (в 45 км к северо-западу от Блумфонтейна (Bloemfontein), озеро Хагенштедт). Культурные комплексы включают ранний MSA, датированный 279±47 тыс. лет назад, сильно ретушированную форму MSA возрастом 157±21 тыс. лет назад и минимально ретушированный комплекс MSA из ряда заселённых горизонтов возрастом 121±6 тыс. лет назад. Здесь обнаружен череп раннего Homo sapiens, датированный 259±35 тыс. лет назад (электрон-спин-резонанс на человеческом зубе, ассоциированном с черепом) [25; 26]. Scerri et al. (2018) рассматривают Флорисбад как представителя одной из линий ранних Homo sapiens, сформировавших в результате гибридизации и эволюции человека современного анатомического типа [38].
Брокен-Хилл, Кабве / Broken Hill, Kabwe (Родезийский человек; Rhodesian Man) — череп гейдельбергского человека (Homo heidelbergensis), найденный в Юго-Западной Замбии (в прошлом — Северная Родезия) (г. Кабве, близ станции Брокен-Хилл, свинцово-цинковый рудник Брокен-Хилл). По данным датирования методом урановых рядов имеет возраст 299±25 тыс. лет [19].
Рабат — верхняя челюсть с зубами. Обнаружена в Марокко, южнее г. Рабата, в карьере Кебибат, в литологическом слое S2, для которого получена дата 137±7 тыс. лет назад. Исследователи полагают, что этот слой был сформирован между концом MIS 7 и, вероятно, концом MIS 6. Останки человеческого черепа, найденные в Кебибате, принадлежат прибрежной дюне, образовавшейся во время MIS 6. С палеоантропологической и геохронологической точки зрения останки человека из Кебибата характеризуются более архаичной морфологией и являются более древними, чем другие останки человека, найденные в прибрежных пещерах Темары). Более того, сравнение внешнего вида черепов человека со стоянки Кебибат и стоянки Джебель-Ирхуд позволяет предположить, что морфологически в стоянке Джебель-Ирхуд черепа менее архаичны и относительно более древние, их возраст составляет 300 тыс. лет [15].
Согласно результатам последних исследований Ирхуд 1 — Homo heidelbergensis [3, с.105], а археологические слои в Джебель Ирхуде датируются от 240±35 до 378±30 тыс. лет назад, в среднем 315±34 тыс. лет назад (термолюминесцентные даты). Непосредственно на останках ребёнка Ирхуд 3 получена дата 286±32 тыс. лет назад [24].
Хоупфилд (Салданья (Saldanha, Hopefield 1, Elandsfontein)) — черепная коробка мужчины найдена в Эландсфонтейне, Салданья (Южная Африка). Предполагаемый возраст: 150–400 тыс. лет (?). Точная датировка слоёв местонахождения Салданья невозможна. Сопутствующие орудия относятся к нескольким культурным традициям: финальному ашелю, среднему каменному веку и позднему каменному веку. Таксономическое определение: Homo erectus soloensis (нгандонгский, или солойский человеr) или Homo heidelbergensis. Исследователи отмечают похожесть этого черепа на Брокен-Хилл 1 (Кабве) [11].
Сале, Эль Хамра (El Hamra) — предполагаемый возраст 400 тыс. лет назад [36].
Обнаруженные в пещере Thomas Quarry I окаменелости гомининов, явно близки к предковой линии Homo sapiens [33].
Нгандонг, местонахождение ископаемого человека (явантроп; от 550 до 25 тыс. лет назад) на о. Ява (Индонезия). На левом берегу р. Соло (=Кали-Бенгаван) близ села Нгандонг обнаружены черепные крышки и берцовые кости 12 человек: 8 взрослых (мужчины: Соло 5, 9, 10, 11; женщины: Соло 1, 4, 6, 8), 1 ребёнка мужского пола (Соло 2) и 3 — неопределённого возраста. Почти у всех черепов было проломлено основание (вероятно, признак каннибализма). Индивид Нгандонг V (он же Соло 4) и три других были убиты ударом по голове. Найдены также грубые каменные отщепы, пластины, каменный шар; костяные нож с отполированным краем и гарпун, остроконечное орудие из оленьего рога. Возможно, орудия и фауна с людьми из разных слоёв. Мозаика эректоидных и сапиентных черт, сходство с черепом из Кабве позволяют отнести его к гейдельбергскому человеку, а поздний возраст заставляет считать его реликтовой популяцией этого вида. Некоторые зарубежные исследователи рассматривают гоминид из Нгандонга как предков современных австралийских аборигенов [9]. Hawks et al. (2000), провели поиск потенциального предка WLH-50 среди трёх популяций (Нгандонг, африканцы позднего плейстоцена, гоминиды Леванта из Схула и Кафзеха). В результате они пришли к выводу, что гоминиды Нгандонг или подобная им популяция, наряду с двумя другими группами, внесли значительный вклад в родословную WLH-50 и современных австралийцев-аборигенов [21].
KNM-ES 11693 или Eliye Springs (Элие-Спрингс) — череп взрослого человека, найденный на северо-западе Кении (деревня Элие-Спрингс на западном берегу озера Туркана). Эта находка не имеет чёткого стратиграфического положения и датируется ориентировочно 300–200 тыс. лет назад. На сайте Смитсоновского Национального музея естественной истории (Вашингтон, США) определён как Homo heidelbergensis [18]. Некоторые исследователи относят его в промежуточный между гейдельбергским человеком и Homo sapiens вид Homo helmei [12].
Ндуту 1 (Ndutu 1) — череп, найденный вблизи озера Ндуту (Танзания, юго-восточная часть плато Серенгети, близ вулканического кратера Нгоронгоро, около западного конца Олдувайского ущелья). Форма реконструированного свода черепа Ндуту оказалась наиболее близкой к Sima de los Huesos (SH) 5, в то время как его сагиттальный профиль был наиболее похож на профиль Кабве. Предполагаемый возраст — около 450 тыс. лет назад [30].
Омо 2 (Omo II) — черепная коробка, найденная в формации Кибиш, в Эфиопии (Kibish formation, Ethiopia), по общей форме напоминающая черепа из Нгандонга. Предполагаемый возраст: 104-195-196 тыс. лет назад (⁴⁰Ar/³ꝰAr; 2008 год) [29].
Часть верхней челюсти и практически неповреждённый свод черепа человека Нгалоба (Ngaloba) LH 18 найден на местонахождении Нгалоба (Ngaloba) в местности Лаэтоли (Laetoli), вблизи озера Эяси (Lake Eyasi), в 50 км южнее Олдувая, в Танзании. Слои Нгалоба в которых он был обнаружен датируются примерно 120±30 тыс. лет назад [28]. На сайте Смитсоновского Национального музея естественной истории (Вашингтон, США) он отнесён к виду Homo sapiens, т.е. к человеку современного анатомического типа и датирован — около 120 тыс. лет [32]. Magori, Day (1983), изучавшие череп Ngaloba LH18 отметили наличие на нём смеси как архаичных, так и современных особенностей (например, толстые надглазничные торы (supraorbital tori), которые тем не менее чётко разделены на медиальный и латеральный отделы, общая ширина черепа и изогнутая затылочная часть). Объём черепа около 1200 см³. В морфологии LH 18 есть некоторые различия от черепов других ранних африканских Homo sapiens, но достаточно сходств, свидетельствующих о принадлежности их людям одного вида. LH 18 совершенно не похож на изученные черепа человека прямоходящего и неандертальца. Метрический анализ подтверждает связи между LH 18 и другими архаичными черепами Homo sapiens из Африки, особенно Эяси и Омо 1 [28]. По мнению Lahr et al. (2016) присутствие на черепе LH 18 как архаичных, так и современных особенностей может указывать либо на большую морфологическую изменчивость раннего человеческого населения Восточной Африки или на существование нескольких типичных групп в регионе в то время [27]. В исследовании Mounier, Lahr (2019) восточноафриканский LH18 продемонстрировал наименьшее сходство с современными людьми. Североафриканский Ирхуд-1 занял промежуточное положение между современными людьми и неандертальцами, а южноафриканский Флорисбад и, в меньшей степени, восточноафриканские KNM-ES 11693 и Omo II, показали наибольшую близость к современным людям [31].
Омо 1 из формации Кибиш в Эфиопии, возрастом 232±22 тыс. лет назад, считается древнейшим известным человеком современного анатомического типа [41].
II. Предполагаемые Y-хромосомный «Адам» и носители Y-хромосомных гаплогрупп А00 и А0-Т и их прародина
2.1. Y-хромосомный «Адам»
По данным генетиков, всё современное человечество является потомком Y-хросомного «Адама» (ROOT (Y-Chromosome «Adam»)), предположительно жившего в Африке и возраст которого пока точно не определён (рис. 4). Нам известно, что расчёты с помощью программного обеспечение BEAST2 показали, что время жизни последнего общего предка современных людей и неандертальцев (TMRCA) ~408 тыс. лет назад [35]. Соответственно, возраст Y-хросомного «Адама» ~408 тыс. лет назад и судя по имеющимся палеонтологическим находкам он вряд ли был похож на современного человека.
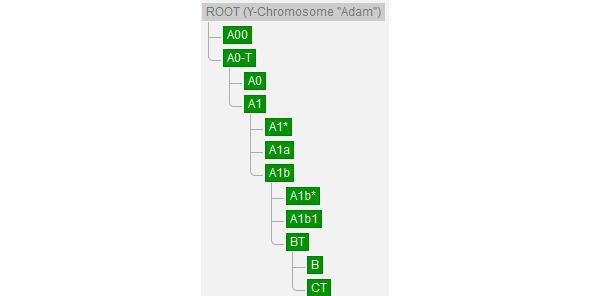
Рис. 4. «Основание» генеалогического древа современного человечества [77]
В настоящее время на Земле проживают потомки двух условных «сыновей» ROOT — A00 (гаплогруппа сформировалась 235900 (243700—228300) лет назад, время жизни последнего общего предка современных её представителей (TMRCA) 37600 (42200—33100) лет назад [78]. Представитель базальной линии этой гаплогруппы, возрастом 7885 лет назад (I10871), обнаружен на северо-западе Камеруна (археологический памятник Шум-Лака (Shum Laka)) и все известные ныне живущие представители патрилинейной наследственности A00 живут в Камеруне [78]. Подробнее о YDNA A00 изложено в параграфе, посвящённом этой гаплогруппе.
Потомками другого «сына» ROOT — A0-T (гаплогруппа сформировалась 235900 (243700—228300) лет назад, время жизни последнего общего предка современных её представителей (TMRCA) 161300 (169900—152900) лет назад), является вся остальная мужская половина современного человечества [79].
Если ориентироваться на дату ~408 тыс. лет назад (время жизни последнего общего предка современных людей и неандертальцев (TMRCA) ~408 тыс. лет назад [35]) и возраст гаплогрупп А00 и А0Т 235900 (243700—228300) лет назад [79], то можно полагать, что на протяжении 173 тыс. лет наши предки представляли единую популяцию, которая разделилась примерно 235 тыс. лет назад.
Для того, чтобы удостовериться, что изначально предки современного человечества составляли единую популяцию, можно сравнить возраст Y-хромосомных и митохондриальных гаплогрупп, представленных у современного человечества. При этом, нужно заметить, что в литературе информация о возрасте последних очень противоречива. Так, например, на сайте компании YFull на филогенетическом древе возраст митохондриальных гаплогрупп не рассчитан (рис. 5).

Рис. 5. «Основание» филогенетического древа мтДНК современного человечества (начинается с гаплогруппы L); HD — это неандертальцы. Носитель базальной линии этой гаплогруппы — неандерталец[1] Sima de los Huesos из Испании, возрастом 394930 (149930—649930) лет назад; DN — денисовцы; NA — неандертальцы. Носитель бальной линии этой гаплогруппы неандерталец Hohlenstein-Stadel из Германии, возрастом 1237300 (61943—182490) лет назад [76]
Schlebusch et al. (2020) приводят для L возраст 179,03 тыс. лет назад, её дочерних клад L0 — 144,931 тыс. лет назад и L1’2’3’4 152,809 тыс. лет назад [61, Fig. S5.7]. У Maier et al. (2022) возраста мтДНК L — нет (у них она обозначена L0'1'2'3'4'5'6'7), но есть время жизни последнего общего предка современных представителей этой гаплогруппы — 143454 (160228—127140) лет назад [48, Table S3].
Можно ориентироваться на расчёты Ingman et al. (2000), согласно которым TMRCA для мтДНК составляет 171500±50000 лет назад[2] [42, p.712]. Дата 171500 лет назад очень близка к времени жизни последнего общего предка современных представителей Y-хромосомной гаплогруппы A0-T (TMRCA) -161300 (169900—152900) лет назад) согласно YFull [79]. Если же мы проведём небольшой подсчёт — 171000+50000=221000 лет, то получим цифру близкую к возрасту Y-хромосомных гаплогрупп А00 и А0-Т — 235900 (243700—228300) лет назад [78; 79].
Также можно обратить внимание на время жизни последнего общего предка современных представителей Y-хромосомной гаплогруппы A0-T (TMRCA) 161300 (169900—152900) лет назад) согласно YFull [79], которое примерно совпадает со временем жизни последнего общего предка современных представителей митохондриальной гаплогруппы L0'1'2'3'4'5'6'7 по Maier et al. (2022) — 143454 (160228—127140) лет назад [48, Table S3] и возрастом клад L0 — 144,931 тыс. лет назад и L1’2’3’4 152,809 тыс. лет назад по Schlebusch et al. (2020) [61, Fig. S5.7].
Поэтому можно полагать, что наши предки, Y-хромосомный «Адам» и митохондриальная «Ева» были представителями единой популяции древних людей, обитавшей на небольшой по площади территории.
Установлено, что клады митохондриальной гаплогруппы L достигают своего пикового разнообразия в странах Восточноафриканской рифтовой долины — Эфиопии, Кении и Танзании (рис. 6) [48, Fig. 4]. Данный факт, по-видимому, является свидетельством формирования человека современного анатомического типа в этом регионе.
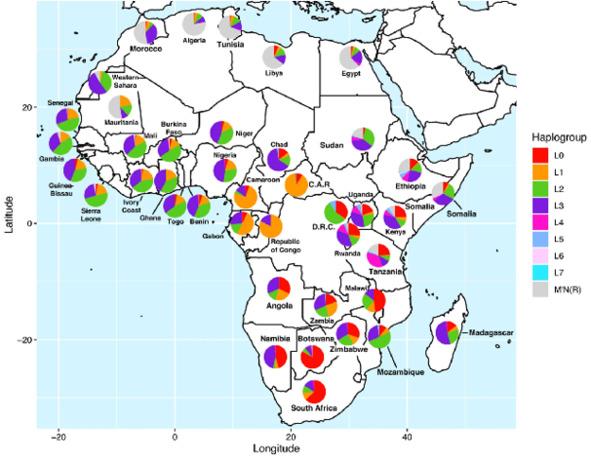
Рис. 6. Частоты распространения различных клад митохондриальной гаплогруппы L в странах Африки (L0 обозначена красным цветом) [48, Fig. 4]
2.2. Археологические культуры, представители которых могут претендовать на роль Y-хромосомного «Адама»
Выше мы пришли к выводу, что наиболее вероятной родиной человека современного анатомического типа являются страны Восточноафриканской рифтовой долины — Эфиопия, Кения и Танзания. Однако исследователи называют людьми современного анатомического типа и гоминин из других регионов, например, из Северной и Южной Африки. Поэтому кратко рассмотрим и эти версии.
2.2.1. Джебель-Ирхуд
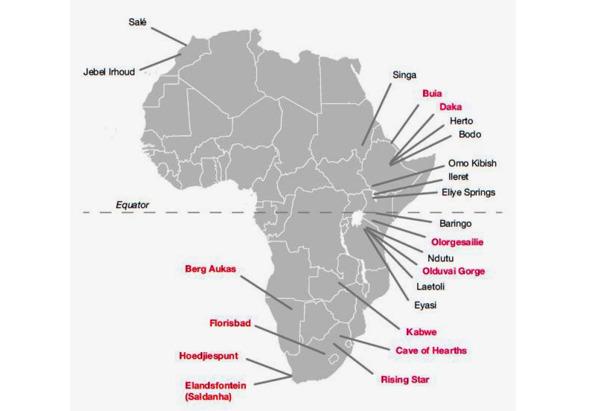
Рис. 7. Местонахождения окаменелостей ископаемых гоминид среднего и раннего плейстоцена в Африке [22]
Пещера Джебель-Ирхуд находится вблизи одноименной деревни, примерно в 100 км к западу от города Марракеш в Марокко и в 75 км от побережья (рис. 7). Она служила рудником для добычи барита (сульфата бария).
Кларк (1977) относит Джебель Ирхуд к мустье[3] [13, с. 109]. По его мнению, мустье в Северной Африке появилось извне. Он выделяет три варианта этой традиции: один (или более) в Нубии[4] — как с бифасами, так и без них, в Киренаике[5] и Магрибе[6]. «Указанные варианты отличаются разными способами техники скола: леваллуа и обработкой дисковидных нуклеусов. Доминирует техника леваллуа, поэтому эти североафриканские археологические местонахождения называются иногда „леваллуа-мустьерскими“» [13, с. 107]. Кларк (1977) пишет: «Указанные мустьерские коллекции представляют уже вполне развитую археологическую традицию, и в настоящее время в нашем распоряжении имеется мало доказательств её появления в Северной Африке в результате эволюции ашельских орудий. Это даёт возможность предполагать, что мустьерская традиция появилась в этих районах либо в результате диффузии, либо была принесена мигрантами с другого континента. Вариант из Киренаики имеет много общих черт с вариантом из Палестины, однако оба они отличаются от нубийского варианта, для которого типично большое количество зазубренных орудий, а также от магрибского варианта, который, в свою очередь, демонстрирует своеобразную региональную специфику. В это время в Северной Африке появляются некоторые евро-азиатские виды животных — медведь, европейский носорог и олень — и с ними, возможно, появляется неандертальский человек, производитель мустьерских орудий» [13, с. 109].
Hublin (2000) отмечает, что мустьерская индустрия, представленная в пещере Джебель Ирхуд, напоминает тип Ла-Феррасси (La Ferrassie) на юго-западе Франции [39, p.159], которая создана неандертальцами. В пещере Ла-Ферасси обнаружены останки трёх неандертальцев, для которых установлен возраст: погребение неандертальца Ла-Ферраси-1 (рис. 8) относится ко времени между 51,8 и 32,1 тыс. лет назад, неандерталки Ла-Ферраси-2 — 49,6—44,1 тыс. лет назад, ребёнка Ла-Ферраси-8 — 44,9—39,9 тыс. лет назад [34].
Необходимо также отметить, что Hershkovitz et al. (2018) говорят о сходстве индустрий Джебель Ирхуда и Табун D из Леванта [38].

Рис. 8. Homo neanderthalensis Ля-Ферраси 1. Мужчина 45—50 лет. Место находки: Франция, департамент Дордонь, пещера Ля-Ферраси. Датирован: 51,8 и 32,1 тыс. лет назад. Слева портрет, выполненный 3D-художником Philip Edwin [45], справа реконструкция из сети Х (пользователь InterspeciesFace) [32]
В пещере Джебель-Ирхуд обнаружены останки фауны, неатерийские археологические находки MSA (non-Aterian Middle Palaeolithic/Middle Stone Age (MSA) industries) и останки нескольких ископаемых гомининов. Ископаемые человеческие останки относятся к низшим слоям стратиграфической последовательности, наиболее известными из которых являются череп древнего человека (известен как Ирхуд 1, JI1) (рис. 9), черепная коробка взрослого индивида Ирхуд 2 (JI2) и нижняя челюсть ребёнка Ирхуд 3 (JI3) [65].
Археологические слои в Джебель Ирхуде датируются от 240±35 до 378±30 тыс. лет назад, в среднем 315±34 тыс. лет назад (термолюминесцентные даты). Непосредственно на останках ребёнка Ирхуд 3 получена дата 286±32 тыс. лет назад [41].

Рис. 9. Джебель Ирхуд 1. Портрет, выполненный 3D-дизайнером Cícero Moraes [20]
Stringer (2016) констатирует, что Ирхуд 1 и Ирхуд 2 демонстрируют некоторое фенетическое сходство с людьми из Кафзех (Qafzeh), Схула (Skhul) и Херто (Herto), хотя у них отсутствует расширение верхней теменной области. По форме свода черепа (но не лица) имеется общее сходство с окаменелостями Сима (Sima fossils) и другими ранними неандертальцами [65]. По мнению Stringer (2016), Ирхуд 1 не является анатомически современным Homo sapiens, хотя имеет намёки на «современный» базикраниальный угол изгиба в соотношении основания черепа и лица («modern’ basicranial flexion in the relationship of the face and vault) [65].
В отличие от Stringer (2016), Hublin (1992) считает обитателей Джебель Ирхуда предшественниками создателей атерийской индустрии и людьми современного анатомического типа. По его мнению, явные синапоморфии[7] с современным человеком в сочетании с некоторыми сохранившимися плезиоморфными[8] признаками указывают на несколько более примитивный (и более древний?) уровень, чем образцы Кафзех-Схул из Юго-Западной Азии [40].
Hublin et al. (2017) установили, что по строению лицевой части черепа ирхудцы почти не отличаются от современных людей. Однако резко отличаются от нас по форме затылка и внутреннему рельефу мозгового отдела черепа, приближаясь к Homo heidelbergensis и Homo erectus — он удлинён в переднезаднем направлении и не имеет выраженной «глобулярной» формы, характерной для современных людей. На черепе Ирхуд 1 выражен так называемый «шиньон» — вариант строения затылочной кости с резким перегибом между её верхней и нижней частями, встречающийся у архантропов и иногда у неандертальцев [41;60].
Исследователи объясняют подобные черты обитателей Джебель Ирхуда тем, что в ходе эволюции у людей сначала появились современные черты лица, а лишь затем — нынешняя форма мозга. Это предполагает, что форма мозга и его функции (когнитивные способности) развивались в рамках линии Homo sapiens [41; 60]. Они делают вывод, что мозаика особенностей, включающая морфологию лица, нижней челюсти и зубов, соотносящая людей из Джебель-Ирхуд с ранними или недавними анатомически современными людьми и более примитивная нейрокраниальная и эндокраниальная морфология, в сочетании с возрастом 315±34 тыс. лет, делают Джебель-Ирхуд старейшим местом обитания гоминидов в среднепалеолитической Африке, документирующим ранние стадии Homo sapiens. Это показывает, по их мнению, что эволюционные процессы, лежащие в основе появления человека современного анатомического типа, охватывали весь африканский континент, т.е. эти древние люди были частью обширной популяции, занимавшей всю территорию среднепалеолитической Африки [41;60].
Бахолдина (2022) не согласна с Hublin et al. (2017). Она констатирует, что «Сочетание развитого надглазничного рельефа, низкого свода и резкого перегиба затылка в комбинации с наличием клыковыми ямками позволяет диагностировать Ирхуд 1 как Homo heidelbergensis,…» [1, с. 105]. Палеонтологи Джеффри Шварц (Jeffrey Schwartz), Мария Мартинон-Торрес (María Martínon-Torres) также не согласны с тем, что останки из Джебель-Ирхуд принадлежат виду Homo sapiens, поскольку, как заметила Мартинон-Торрес, у их черепов не выражен подбородок и лоб так, как они выражены у современного человека [25].
В исследовании Mounier, Lahr (2019) Ирхуд-1 занял промежуточное положение между современными людьми и неандертальцами. Они после моделирования филогенетических связей между древними и современными черепами, выдвинули два предположения:
1) окаменелости из Марокко, по-видимому, представляют собой останки популяции, которая в среднем палеолите смешивалась с неандертальцами и не внесла вклада в появление современных людей. Основано данное предположение на том, что Ирхуд-1 показывает морфологическую близость с неандертальцами. У Ирхуда-11 нет подбородка, характерного для современного человека и имеющегося у юного Ирхуда-3;
2) альтернативная гипотеза относительно североафриканской популяции, представленной окаменелостями из Джебель-Ирхуда, может основываться на результатах геномного анализа, предполагающих, что митохондриальная ДНК (мтДНК) предков неандертальцев, общая с мтДНК окаменелостей Денисова и Сима-де-лос-Уэсос (Sima de los Huesos), была заменена африканской мтДНК между 460 и 219 тыс. лет назад. Учитывая фенотипическое сходство Ирхуд-1 как с неандертальцами, так и с ранними Homo sapiens, возможно, что окаменелости Ирхуда представляют собой местных потомков африканской популяции, которая расселилась из Африки во время событий в Зелёной Сахаре, связанных с MIS 9, и, таким образом, связаны с африканскими популяциями, которые в это время мигрировали в Европу и, следовательно, внесли свой вклад в эволюцию «классических» неандертальцев [56].
Ближе всего к истине, на мой взгляд, Mounier et al. (2019), которые считают, что Ирхуд-1 — представитель популяции Homo heidelbergensis, основная часть которой мигрировала в Европу и стала предком неандертальцев и Бахолдина (2022), согласно которой, Ирхуд 1 — это Homo heidelbergensis и соответственно, люди из пещеры Джебель-Ирхуд, относятся к этому виду древних людей.
2.2.2. Форсмит (Fauresmith)
Существует гипотеза, что прародина человека анатомически современного типа находится в Южной Африке, к югу от реки Замбези, на севере современной Ботсваны [28]. Поэтому рассмотрим индустрии MSA Южной Африки.
Форсмит (Fauresmith) — региональная южноафриканская переходная индустрия от раннего к среднему палеолиту. Индустрия/культура получила своё название по находкам близ г. Фаурсмит (Форсмит) в ЮАР. Стоянки типа Форсмит найдены также на территории Восточной Африки вплоть до Эфиопии [18].
Clark (1986) датировал форсмит в регионе Оранжевой реки между 115—80 тыс. лет до н. э. [29, p.287].
Kuman et al. (2020) пишут, что форсмит датировался 500—280 тыс. лет назад, то есть от позднего ашеля до самой ранней даты появления MSA на юге Африки, в Флорисбаде (Florisbad) — 279±47 тыс. лет назад (ЭПР-дата). Даты со стоянки Кату Пан, полученные на основании комбинации результатов OSL-датирования, U-серий и электронного спинового резонанса, показали, что индустрия форсмит древнее и появилась примерно 0,6—0,5 млн. лет назад[9]. Возраст самых поздних комплексов индустрии форсмит в пещере Вандервер (Wonderwerk Cave) имеет сомнительную дату 187±8 тыс. лет назад, полученную на основе U-series датировки фррагмента сталагмита [44]. По мнению Kuman et al. (2020) данная индустрия развивается в результате результате изменений в поведении, связанных с развитием Homo sapiens [44].
Для индустрии форсмит характерно сочетание ашельской и леваллуазской техники: рубила, кливеры, скребки, скрёбла довольно часто встречаются, сочетаясь с леваллуазскими нуклеусами, пластинами, ретушированными остриями и орудиями геометрических форм [3].
Наиболее изученные стоянки с индустрией форсмит — Кату Пан и Кантин-Копье (Canteen Kopje)[10], расположенные в Северо-Капской провинции ЮАР.
2.2.2.1. Кату Пан
Археологический комплекс Кату Пан представляет собой скопление нескольких археологических местонахождений, относящихся в основном к каменному веку, расположенных в окрестностях города Кату, на южной окраине пустыни Калахари, в Северо-Капской провинции ЮАР.
Кату-Пан-1
В Кату-Пан-1 (КП1) (слой 4b), обнаружена индустрия раннего палеолита. Выше него, в слое 4а, датируемом комбинацией OSL и ESR/U-серий, примерно 500 тыс. лет назад, содержится индустрия форсмит (Fauresmith), относящаяся к рубежу между ранним и средним палеолитом, которая характеризуется отщеповой технологией, изготовлением пластин и односторонних ретушированных острий, которые, по мнению археологов, представляют собой самые ранние наконечники копий. Применялись преференциальная и рекуррентная левалуазские стратегии расщепления и нелеваллуазские стратегии [72; 73; 74].
Пластинчатая индустрия форсмит местонахождения Кату Пан 1 (слой 4a), является наиболее ранней в Южной Африке. Культуросодержащий слой, в котором она обнаружена, имеет даты: ОСЛ — 464±47 тыс. лет назад. Для зуба зебры, извлечённого из этого слоя, комбинированным методом урановых серий и электрон-спин-резонанса (ESR) получены даты 542±140/107 тыс. лет назад [58].
Для вышележащего слоя 3 (относимого к среднему каменному веку) методом OSL был получен возраст 291±45 тыс. лет [72; 73; 74].
Кату-Пан-6
Местонахождение под открытым небом Кату Пан 6 (КП6) расположено примерно в 500 м к юго-востоку от КП1. В Кату Пан 6 выявлена сложная последовательность четырёх каменных индустрий, включающая ассамбляжи форсмита — слой 12 (соответствует слою 4а KП1, датируемому ~500 тыс. лет назад), раннего среднего палеолита (MSA) — слой 11 (ранний тип MSA; сопоставим с индустрией слоя 3 KП1, датируемого 291±45 тыс. лет), среднего палеолита (комплекс Ховисонс-Порт (Howiesons Poort) (HP)) — слои 9—10 и позднего палеолита ((LSA) — верхние слои; комплекс Уилтон (Wilton) с микролитическим компонентом, подобным, обнаруженному в слоях 3b–4c пещеры Вондерверк (Wonderwerk)). В Кату Пан 6 слои 7 и 8 археологически стерильны [47]. Индустрия позднего палеолита Уилтон (the Later Stone Age Wilton industry) в KП6, датируется между 5,7±0,3 тыс. лет назад и 2,3±0,1 тыс. лет назад [47].
Lukich et al. (2019) возраст раннего среднего палеолита в слоях 11 и 12 (KП6—2 и KП6—1 соответственно) определили в 120—95 тыс. лет, что значительно моложе возраста 291±45 тыс. лет для аналогичной индустрии из соседнего местонахождения KП1, датированной в 2010 году [47]. Пытаясь объяснить такую огромную разницу в датировках, Lukich et al. (2019) отмечают, что ранний каменный материал MSA как из KП1, так и из KП6 сохранился очень плохо и не был предметом детального анализа, что оставляет открытой возможность того, что это разные индустрии, которые подверглись сходным тафономическим процессам. Точно такая же ситуация с датировкой индустрии форсмит (Fauresmit) из слоя 12 KП6, ~155 тыс. лет назад, что намного моложе, чем ожидалось (возраст ассамбляжа форсмит в KП1, ~500 тыс. лет назад) [47].
2.2.2.2. Истоки индустрии MSA в Южной Африке и таксономический статус её создателей
Даты индустрии форсмит Южной Африки имеют огромный разброс дат от 542±140/107 тыс. лет назад / 464±47 тыс. лет назад (пластинчатая индустрия форсмит местонахождения Кату Пан 1 (слой 4a)) [58] до ~155 тыс. лет назад (слой 12 Кату-Пан-6, соответствующий слою 4а Кату Пан 1) [47].
Поэтому можно выдвинуть несколько предположений.
1) Наиболее вероятно, что более точны даты, полученные Lukich et al. (2019) в 2019 году [47].
Если исходить из дат Lukich et al. (2019) можно обратить внимание, что дата 155 тыс. лет назад для слоя 12 КП6 (соответствует слою 4а KП1 с индустрией форсмит, датируемому ~500 тыс. лет назад) близка к возрасту предковой для большинства современного человечества Y-хромосомной гаплогруппы А1 (сформировалась 161300 (169900—152900) лет назад, TMRCA 133400 (139000—127800) лет назад) и А0 (сформировалась 161300 (169900—152900) лет назад), встречающейся только в Африке [79]. Это может означать, что создатели индустрии форсмит были ветвью единой для современного человечества популяции, мигрировавшей с северо-запада Африки на юг этого континента и впоследствии, вымершей.
2) Если допустить, что верны даты, полученные в 2010 году Porat et al. (2010), то вполне возможно, что индустрия форсмит появилась в Южной Африке вместе с Homo erectus с северо-востока этого континента. В то же время, если правы Hershkovitz et al (2018), которые утверждают, что древнейшая индустрия Кату-Пан, имеет сходство с индустрией в Джебель-Ирхуде в Магрибе [38], гоминин из которого, скорее всего, Homo heidelbergensis [1, с. 105], то и создатели южноафриканской индустрии в Кату-Пан, должны относиться к этому виду древних людей.
Подтверждением того, что создатели индустрий MSA пришли в Южную Африку с севера, служит тот факт, что в Восточной Африке переход от ашеля к MSA произошёл до 284 тыс. лет назад в формации Каптурин в Кении. В той же Кении, в Олоргесайли (Olorgesailie), комплексы, датируемые около 615—499 тыс. лет назад, имеют черты, отличающие их от более старых ашельских находок на том же участке и более характерные для MSA, такие как небольшие нуклеусы (small cores) (с некоторых из них были сняты несколько пластинок), мелкие отщепы и более экзотические источники камня, или акцент на одном источнике камня более высокого качества. Переход к MSA, в котором отсутствуют ашельские элементы, датируется на этом местонахождении 305—320 тыс. лет назад [44].
Древнейшая дата для MSA Южной Африки — 279±47 тыс. лет назад происходит с местонахождения Флорисбад [3]. Гоминид Флорисбад (Florisbad), живший здесь 259±35 тыс. лет назад [56, Table 1], отнесён к виду Homo heidelbergensis [2, с. 84].
В пещере Бордер (Border Cave) на границе Свазилэнда и ЮАР (провинция Квазулу-Наталь) ЭПР-даты для древнейших слоёв MSA указывают на возраст свыше 200 тыс. лет [3]. В пещере 13В в Пиннакл-Пойнт на южном побережье Мыса Доброй Надежды древнейшие слои MSA имеют даты порядка 180 тыс. лет [3], ~164±12 тыс. лет [49]. Здесь найдены окаменелости двух гомининов. Обнаруженный резец метрически занимает промежуточное положение между гомининами среднего плейстоцена и современными людьми. Каменные артефакты типологически и технологически напоминают региональный вариант MSA (индустрия залива Моссель), но значительно меньше [50].
В пещере Вандерверк возраст индустриии MSA, согласно результатам ТУ-датирования, составляет 150–200 тыс. лет [3]. Останки гоминин не известны.
Кроме того, южноафриканская стоянка Дюйнефонтейн 2 (Duinefontein 2), относящаяся к позднему ашельскому периоду, может быть переходной. ОСЛ-даты для песков, подстилающих и перекрывающих верхний ашельский слой этого памятника, говорят о том, что он не древнее 290 и не моложе 270 тыс. лет назад, а фауна соответствует возрастному диапазону 250—300 тыс. лет назад. Однако диагностические ашельские типы орудий опубликованные с этой стоянки, ограничены одним целым и двумя сломанными рубилами, а также несколькими фрагментами рубила, и поэтому ассамбляж может быть переходным [44].
2.2.3. Олоргесайли (Olorgesailie)
Открытое местонахождение Олоргесайли расположено между двумя потухшими вулканами примерно в 70 км от Найроби на юге Кении, в восточной части Рифтовой долины (рис. 7). Названо в честь горы Олоргесайли, расположенной неподалёку. Здесь, вместе с ашельскими рубилами, обнаружены фрагменты свода черепа Homo erectus, названного KNM-OL 45500, датируемые 900—970 тыс. лет (40Ar/39Ar, магнитостратиграфия, стратиграфия) [33]. Отмечается близость KNM-OL 45500 по размерам к образцам из Дманиси (Грузия) и Homo naledi [55]. Внешность Homo erectus представлена на рис. 10).
Первые свидетельства существования человека на этой территории датируются 1,2 млн. лет назад. Исследование показало, что обитатели Олоргесайли изготавливали на протяжении 700 тыс. лет из местного сырья рубила. 305—320 тыс. лет назад у них произошёл резкий сдвиг в поведении и стратегиях выживания, появились новые технологии изготовления орудий, признаки обмена с другими группами людей и использование в быту цветных пигментов. В частности, орудия, в отличие от универсальных рубил, стали более специализированными. Многие из них представляла собой наконечники, предназначенные для прикрепления к древку и, по-видимому, использовавшиеся в качестве метательного оружия. Часть орудий имела форму скребков или шил. Небольшие каменные наконечники были изготовлены из обсидиана, источники которых находились на расстоянии 24—88 км (15—55 миль) от Олоргесайли, где нет собственного источника этого камня. Это указывает на то, что между различными группами людей в Африке мог существовать обмен. Были также обнаружены чёрные и красные камни — марганец и охра, а также свидетельства того, что камни были обработаны для использования в качестве красящего материала.

Рис. 10. Скульптура женщины Homo erectus, выставленная в Зале происхождения человека Смитсоновского национального музея естественной истории (Вашингтон, округ Колумбия, США) (реконструкция J.Gurche; фотограф T. Evanson) [57]
Проанализировав образцы горной породы, исследователи установили причины этих поведенческих, технологических и культурных сдвигов у обитателей Олоргесайли. Геологические, геохимические, палеоботанические и фаунистические данные показали, что около 360 тыс. лет назад в регионе начался длительный период нестабильности климата, землетрясения постоянно меняли ландшафт и соответвенно, менялись флора и фауна. Всё это привело к слому привычного образа жизни древних людей. Поэтому они были вынуждены приспосабливаться к непредсказуемым условиям жизни. Технологические инновации, сети социального обмена и ранняя символическая коммуникация стали теми факторами, которые помогли древним людям выжить и получить необходимые им ресурсы [24; 30; 59].
2.2.4. Каптурин

Рис. 11. Местоположение формации Каптурин и других местонахождений MSA [21]
Формация Каптурин (Kapthurin Formation) находится на холмах Туген (Tugen Hills) в Кенийской рифтовой долине (Kenya Rift Valley) (рис. 11) и представляет собой серию осадочных и вулканических отложений среднего и позднего плейстоцена толщиной 125 м, обнажившихся на площади более 150 км² к западу от озера Баринго. Формация названа в честь врезающейся в неё реки Каптурин. Слои тефры[11] были датированы, пронумерованы снизу доверху от K1 до K5 (рис. 12) и коррелированы, чтобы обеспечить надёжную хронологию горизонтов Каптурина. В ней сохранилось множество археологических и палеонтологических памятников [31; 43].
В формации Каптурин десять археологических памятников, среди которых GnJh-41 GnJh-42 и GnJh-57, находятся в слоях, датированных радиометрическим методом (⁴⁰Ar/³ꝰAr) ~0,55–0,52 млн. лет назад (545±3 и 509±9 тыс. лет назад) (рис. 12). Их ассамбляж представлен отщепами, изготовленными из небольших галек (informal flake industry made on small cobbles). Ручные рубила (handaxes) редки, составляя лишь 1,1% или даже 0,5% от общего числа артефактов. Редкость рубил примечательна, поскольку в этот период ашельская индустрия, характеризующаяся рубилами, была широко распространена в Восточной Африке [31].
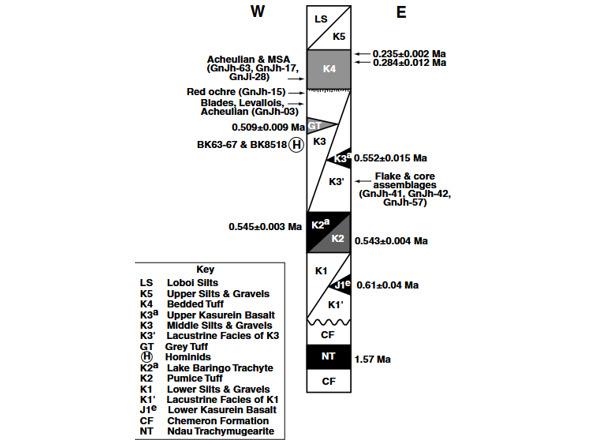
Рис 12. Схематический стратиграфический разрез формации Каптурин [31, Fig. 2].
На стоянках GnJh-42 и GnJh-50 формации Каптурин в культуросодержащих слоях, датированных (⁴⁰Ar/³ꝰAr) 545±3 и 509±9 тыс. лет назад, зафиксировано наиболее ранее в мире появление пластинчатой индустрии[12] [43]. Отметим, что пластины в формации Каптурин появились значительно раньше, чем произошло расхождение Homo neanderthalensis и Homo sapiens [43] (расчётное время жизни последнего общего предка современных людей и неандертальцев (TMRCA) ~408 тыс. лет назад [35]). Оформление нуклеусов и скалывание пластин на этих стоянках типологически близко, по мнению исследователей, к объёмному методу, применявшемуся на стоянке Хуммала[13] в Центральной Сирии [43, p.195].
Johnson, McBrearty (2010) отмечают, что стандартизированные заготовки, обнаруженные на стоянках GnJh-42 и GnJh-50 формации Каптурин, имеют параллельные или почти параллельные стороны и соотношение длины к ширине, равное или превышающее 2:1. Подобные стандартизированные заготовки встречаются в разном количестве на многих европейских поселениях эпохи верхнего палеолита (UP; 40 тыс. лет назад) и африканского позднего каменного века (LSA; 50 тыс. лет назад) [43, p.193]. Если это действительно так, то в обитателях стоянок GnJh-42 и GnJh-50 формации Каптурин, можно видеть древнейших прямых предков современного человечества.
2.2.4.1. Гоминиды формации Каптурин
В формации Каптурин обнаружены останки гоминид KNM-BK 63—67 и KNM-BK 8518 в слоях с датами ~543±9 и 509±4 тыс. лет назад, соответственно. Учитывая их стратиграфическое положение на 0,7 и 3 м ниже пемзового туфа (Pumice Tuff), их возраст составляет 510–512 тыс. лет назад. Эти гоминиды среднего плейстоцена являются ближайшими современниками гоминидов Бодо (датирован методом ⁴⁰Ar/³ꝰAr в интервале 640–550 тыс. лет назад) из Эфиопии, и ОН 11, ОН 23 и Ндуту из ущелья Олдувай в Танзании, возраст, которых оценивается в 490–780 тыс. лет назад [31].
Нижние челюсти KNM-BK-67 и KNM-BK-8518 имеют в целом архаичную морфологию и у них отсутствуют подбородки, что исключает их из Homo sapiens. Правая плюсневая кость KNM-BK-63 и проксимальная фаланга KNM-BK-64 также имеют архаичный внешний вид. KNM-BK-65 демонстрирует как архаичные особенности, так и признаки, которые можно увидеть у Homo sapiens. Таксономически их определяют, как Homo erectus, «архаичный» Homo sapiens, Homo rhodesiensis [31].
Как уже упоминалось, расчёты с помощью программного обеспечение BEAST2 показали, что время жизни последнего общего предка современных людей и неандертальцев (TMRCA) ~408 тыс. лет назад [35]. Гоминиды формации Каптурин, если их возраст определён верно, старше Y-хромосомного «Адама» примерно на 100 тыс. лет. Вполне возможно, что популяция, обитавшая в формации Каптурин в этот период — предковая для людей современного анатомического типа и неандартальцев (и Nesher Ramla Homo из Леванта, датируемого 140–120 тыс. лет назад (пещера Нешер Рамла)[14] и др.). Вероятно, о развитии этой популяции в сторону сапиентации является наличие у KNM-BK-65 как архаичных особенностей, так и признаков, которые можно увидеть у Homo sapiens [31].
На принадлежность популяции древних людей из формации Каптурин к общим предкам современного человека и неандертальцев, а также Nesher Ramla Homo, косвенно указывают археологические данные. Так, например, по мнению Деревянко (2016) первичное пластинчатое расщепление, техника леваллуа (протолеваллуа), из Восточной Африки проникли в Левант, где известны на стоянке Гешер Бенот Яаков и более ранних местонахождениях [5]. Однако комплексы Каптурин «и стоянки Гешер Бенот Яаков имеют мало общего, их невозможно объединить в один комплекс» [5]. Объясняет Деревянко (2016) это следующим образом: «любые технологические традиции во время длительных передвижений изменяются в связи с необходимостью адаптации мигрирующих популяций к новым экологическим условиям (другой климат, животный и растительный мир, исходный материал для изготовления каменных орудий и т.д.), а также под влиянием технологических традиций автохтонного населения» [5, с.16].
Пластинчатая технологии, впервые зафиксированная в Каптурин, получила широкое распространение в Леванте на ашело-ябрудийском этапе позднего ашеля. В ярко выраженной форме пластинчатое расщепление прослеживается в амудийской индустрии на местонахождении Кесем в Израиле (400–200 тыс. лет назад) [5, с. 22].
2.2.4.2. Эволюционное развитие индустрий формации Каптурин в ашельский период. Каптурин — родина технологии центростремительного леваллуа, пластинчатого производства и символической деятельности
Средняя часть формации Каптурин датируется от 0,509±0,009 до 0,284±0,012 млн. лет назад. В отложениях этого возраста (стоянки GnJh-17, GnJh-63 и GnJi-28 [67]) найдены острия/наконечники (points), типичные для среднего каменного века (MSA). Эти даты старше возраста MSA в других частях Восточной Африки на 49 тыс. лет (датируется ~234 тыс. лет назад) и устанавливают возраст перехода от ашеля к MSA в регионе [31]. В этом же интервале встречаются свидетельства использования леваллуазской техники[15] изготовления как мелких отщепов, так и заготовок бифасов, систематического изготовления пластинок (blades)[16], использования и обработки красной охры. Присутствие пластин (blades) и красной охры важно, поскольку пластины означают высокую степень технической компетентности, а красная охра предполагает символическое поведение [31].
Красная охра обнаружена на стоянке GnJh-15, расположенной на береговом склоне р. Бартекечо, притока р. Каптурин и лежит непосредственно под слоем туфа К4, датируемого 0,284±0,012 млн. лет назад. Красная охра с этой стоянки значительно старше охры со среднепалеолитических памятников Южной Африки, таких как пещеры на реке Класье (Klasies River Caves), датированных различными методами до 120 тыс. лет назад, пещера Бломбос, имеющей дату — до 70 тыс. лет назад и Твин-Риверс (Twin Rivers) в Замбии, датированной до ~230 тыс. лет назад [31].
Сравнение больших леваллуазских отщепов, полученных в результате центростремительного расщепления галек из лавы и выработанных из них путём ретуширования ручных рубил (handaxes), кливеров (cleavers) и других подобных орудий со стоянки LHA/GnJh-03 возрастом около 400 тыс. лет назад (395.6±3.5 тыс. лет назад) с находками из четырёх классических ашельских комплексов центральной Рифтовой долины: Кариандуси (Kariandusi)[17], Олоргесайли CL1—1 (Olorgesailie CL1—1)[18], Олоргесайли Пачка 6/7 (Olorgesailie Member 6/7)[19], и Исинья (Isinya)[20] (рис. 13), выполненных на крупных отщепах, показало, что орудия формации Каптурин отличаются своей тонкостью (как и в формации Каптурин, все рубила и кливера на этих стоянках были изготовлены из мелкозернистой вулканической породы). Это достигалось за счёт симметричной центростремительной обработки пологих выпуклостей верхней поверхности нуклеусов Леваллуа, созданием выступающей гранённой платформы и проксимального скоса верхней поверхности. Кроме того, ашельские рубила и кливера формации Каптурин сравнивались с позднеашельскими подобными орудиями из Каламбо-Фоллс (Kalambo Falls)[21], изготовленными из кварцита. Ни в одном из сравнимаемых ассамбляжей нет свидетельств леваллуазской технологии создания рубил и кливеров [62].
Рис. 13. Местоположение Олдувайского ущелья и ашельских комплексов, сравнимаемых с ашельским комплексом формации Каптурин [62, Fig. 1].
Леваллуазская технология из расположенной выше LHA/GnJh-03 стоянки Коимилот (Koimilot (GnJh-74); возраст на основании геохимической корреляции с датированной вулканической тефрой, составляет около 200 тыс. лет [66]; 200—250 тыс. лет [67]) скорее показывает различия в проявлении технологии производства леваллуазских отщепов, чем в типологии[22], что позволяет предположить развитие местной предшествующей ашельской традиции [66; 67], т.е. традиции леваллуа, зародившейся на ашельской стоянке LHA/GnJh-03, возрастом ~400 тыс. лет назад.
Первоначально McBrearty с соавторами (McBrearty et al. (1996), McBrearty, Brooks (2000)) считали, что позднеашельская пластинчатая индустрия на стоянках GnJh 3, 15, 17 формации Каптурин периода около 280 тыс. лет назад, по технологическим показателям не имела преемственной связи с более древней, возрастом свыше 500 тыс. лет [51; 52]. Позже она с соавтором (Tryon, McBrearty (2006) предположила, что различные методы производства леваллуазских отщепов в пластах, датируемых ~200–250 тыс. лет назад на стоянке Коимилот (Koimilot (GnJh-74), могли возникнуть на основе предшествующей ашельской технологии [67]. Ещё позже она с соавтором (Johnson, McBrearty (2010)) пришла к выводу, что пластины из местонахождений GnJh-42 и GnJh-50, датируемых 545—509 тыс. лет назад и GnJh-03, GnJh-17, GnJh-15, датируемых 509—285 тыс. лет назад, принадлежат к одной традиции, которая сохранялась в Восточной Африке на протяжении большей части среднего плейстоцена [43, p. 198]. Johnson, McBrearty (2010) отмечают, что географически распространённый ашельский технокомплекс, как однородная технологическая традиция, оставался устойчивым к изменениям на протяжении всего своего длительного существования, вплоть до замены индустриями MSA. Эти изменения, по-видимому, связаны с расширением поведенческого репертуара гоминин, включившего в себя действия, необходимые для производства пластин, отщепов и наконечников Леваллуа, а также бифасиально отретушированных наконечников, характерных для MSA. Эти технологические инновации, возможно, в какой-то степени повлияли на анатомические изменения, обнаруженные в палеонтологической летописи гомининов [43, p. 198].
2.2.4.3. Сосуществование популяций древних людей разных видов в формации Каптурин
Tryon, McBrearty (2002, 2006) констатируют, что в формации Каптурин хронологический порядок расположения памятников не показывает чёткого однонаправленного перехода от ашельского периода к MSA. Например, на стоянках GnJh-17 и GnJh-63 (датируются 284±12 — 235±2 тыс. лет назад) видна взаимная стратификация характерных для MSA острий/наконечников (points) и ашельских рубил (handaxes). Наконечники (points) и рубила (handaxes) также встречаются на одном и том же археологическом уровне в GnJh-17 и Ророп-Лингопе (Rorop Lingop, GnJi-28) (датируются 284±12 — 235±2 тыс. лет назад). На местонахождении 108 (Loc. 108) обнаружены кливера (cleavers) в слоях, залегающих над верхними базальтовыми туфами, и в совокупности эти наблюдения свидетельствуют о том, что ашельское производство орудий труда сохранялось и после появления наконечников. Исследователи также отмечают, что, возможно, в формации Каптурин ашельские элементы переслаиваются с сангоанскими, MSA и, возможно, с компонентами форсмита (Fauresmith). В отличие от таких местонахождений как Каламбо-Фоллс (Kalambo Falls)[23], Пещера очагов (Cave of Hearths)[24], Исимила (Isimila)[25] или Мугурук (Muguruk), слои формации Каптурин, содержащие ашельские артефакты (рубила (handaxes)) лежат поверх тех, которые содержат артефакты, определяющие Сангоан (кирки (picks)) и MSA (острия/наконечники (points)). Кирки, являющиеся отличительной чертой сангоана, также встречаются в бесспорно ашельских коллекциях и ассамбляжах MSA. Так, например, в местонахождении GnJh-17 на одном уровне обнаружены рубило, кирка и наконечник. Аналогичные находки известны на уровне 7 Гарба III в Мелка-Контуре [67; 68].
По мнению Tryon, McBrearty (2002), остаётся неясным, является ли технологическое разнообразие, наблюдаемое в пределах формации Каптурин, результатом сложной адаптации гоминид к разнообразной палеосреде в пределах одного бассейна или результатом присутствия здесь нескольких видов гоминид [67]. Они предлагают несколько интерпретаций данному явлению. Во-первых, сочетание артефактов на таких стоянках, как GnJh-17, может представлять собой вариации в позднем ашеле или раннем MSA, отражающие адаптацию к среде обитания или видам деятельности. Во-вторых, они могут свидетельствовать о сохранении более древних методов обработки камня в процессе накопления новых технологий в рамках одной традиции. В-третьих, различные классы артефактов могут обозначать отдельные занятия различных групп современных гоминидов, использующих разные технологии. В-четвёртых, они могут представлять собой палимпсест отложений, накопленных с течением времени [67].
Выводы.
1) Датировки появления леваллуазской техники, использования и обработки красной охры (стоянка GnJh-15), острий/наконечников (points), типичных для среднего каменного века (MSA) (стоянки GnJh-17, GnJh-63 и GnJi-28) в Каптурине ~284 тыс. лет назад более ранние, чем появление подобных артефактов в других частях Восточной Африки ~234 тыс. лет назад. При этом, дата ~234 тыс. лет назад близка к возрасту гаплогрупп А00 и А0-Т — 235900 (243700—228300) лет назад [79]. Это позволяет предположить, что в формации Каптурин обитала предковая для всего современного человечества популяция древних людей.
2) На возможность обитания в формации Каптурин предковой для современного человека популяция Homo heidelbergensis/Homo bodoensis 545±3 — 509±9 тыс. лет назад указывает тот факт, что эти древние люди отличались от остальных популяций ашельского периода Африки орудийным набором (редкость рубил [31]). Здесь же, на стоянках GnJh-42 и GnJh-50, датируемых 545±3 — 509±9 тыс. лет назад, зафиксировано наиболее ранее в мире появление пластинчатой индустрии [43]. Впоследствии стандартизированные заготовки, подобные обнаруженным на стоянках GnJh-42 и GnJh-50 формации Каптурин встречаются в разном количестве на многих европейских поселениях эпохи верхнего палеолита (UP; 40 тыс. лет назад) и африканского позднего каменного века (LSA; 50 тыс. лет назад) [43, p.193].
3) Технология изготовления каменных орудий на ряде стоянок формации Каптурин, датируемых 509—285 тыс. лет назад, ~200–250 тыс. лет принадлежит к традиции, датируемой 545—509 тыс. лет назад [43, p. 198; 68], что может быть свидетельством эволюционного развития в данной формации Homo heidelbergensis/Homo bodoensis в Homo sapiens sapiens.
4) Примерно 200 тыс. лет назад в формации Каптурин появилось новое население. Об этом свидетельствует то, что здесь в период MSA, как полагают археологи, можно выделить два среднепалеолитических локуса, датируемые ∼200 тыс. лет назад. Для индустрии локуса 1 характерны отщепы, преференциальная и рекуррентная леваллуазские стратегии, а также стратегии, не связанные с леваллуа. Для индустрии локуса 2 характерны острия и пластины (blades), снятие пластин с нуклеусов, острия и однонаправленная конвергентная леваллуазская стратегия [66; 68; 69]. Археологи также отмечают появление на одном археологическом уровне орудий характерных для ашеля, сангоана, MSA и, возможно, форсмита (Fauresmith) [67; 68].
Вполне возможно, что в этот период в формации Каптурин началось смешение гоминидов разных видов. Есть гипотеза Mounier, Mirazón Lahr (2019), что люди современного типа, расселившиеся по планете и живущие на ней сейчас, получились в результате смешивания популяций их предшественников из Южной, и, возможно, Восточной Африки [56] (см. ниже: Гипотеза Mounier, Lahr (2019)).
5) Приход людей другого вида, по всей вероятности, привёл к их смешению с аборигенами формации Каптурин (см. ниже), а другая часть аборигенов, предположительно, могла мигрировать на север, в Гадемотта/Кулькулетти. Таким образом, из мигрантов в Гадемотта/Кулькулетти могла сформироваться популяция носителей Y-хромосомной гаплогруппы А0-Т, а в Каптурине — популяция носителей Y-хромосомной гаплогруппы А00. Впоследствии представители последней, под давлением потомков А0-Т мигрировали по зоне экваториаль



















