

автордың кітабын онлайн тегін оқу Экологическая физиология сельскохозяйственных культур
Е. И. Кошкин, Г. Г. Гусейнов
Экологическая физиология сельскохозяйственных культур
Учебное пособие
![]()
Информация о книге
УДК 633(075.8)
ББК 42.1я73
К76
Авторы:
Кошкин Е. И., Гусейнов Г. Г.
Рецензенты:
Кузнецов В. В. – доктор биологических наук, профессор, член-корреспондент РАН, ИФР РАН;
Попов С. Я. – доктор биологических наук, профессор кафедры защиты растений РГАУ – МСХА им. К. А. Тимирязева.
В учебном пособии проанализированы структурные и функциональные особенности адаптации и формирования урожая полевых культур под действием биотических и абиотических факторов в преимущественно техногенных системах земледелия. Приведены визуальные симптомы некоторых инфекционных и неинфекционных заболеваний, рассмотрены физиологические нарушения у культурных и сорных растений при обработке пестицидами. Кратко изложены сведения о биохимии иммунитета растений. Особое внимание уделено использованию физиологических показателей в селекции на урожайность и качество урожая растений при стрессе. Показаны возможности использования для повышения толерантности к стрессорам ряда специальных индукторов устойчивости органической и неорганической природы.
Предназначено для студентов агрономических специальностей, обучающихся по программам магистратуры, а также аспирантов, преподавателей, научных сотрудников, работников сельского хозяйства.
УДК 633(075.8)
ББК 42.1я73
© Кошкин Е. И., Гусейнов Г. Г., 2020
© ООО «Проспект», 2020
СПИСОК СОКРАЩЕНИЙ
АБК — абсцизовая кислота
АДГ — алкогольдегидрогеназа
АФК — активные формы кислорода
АО — антиоксиданты
АОС — антиоксидантные системы
АТО — ассимиляционно-транспирационное отношение
АФК — активные формы кислорода
АЦК — 1-аминоциклопропан-1-карбоновая кислота
БТШ — белки теплового шока
ГДГ — глутаматдегидрогеназа
ГДФ — гуанозиндифосфат
ГК — гибберелловая кислота
ГОГАТ — глутаминоксиглутарат-аминотрансфераза
ГС — глутаминсинтетаза
ГТФ — гуанозинтрифосфат
ДАГ — диацилглицерол
ДДВП — дефицит давления водяных паров
2,4-Д — 2,4-дихлорфеноксиуксусная кислота
ДК — дыхательный коэффициент
ИД — интенсивность дыхания
ИЛП — индекс листовой поверхности
ИТ — интенсивность транспирации
ИТФ — инозитолтрифосфат
ИУ — индекс устойчивости
ИУК — индолил-3-уксусная кислота
ИФ — интенсивность фотосинтеза
ИФN — фотосинтетическая эффективность использования азота
ИЭЗ — изоэлектрическая зона цитоплазмы
КВК — кислородвыделяющий комплекс
кДНК — комплементарная ДНК
ЛДГ — лактатдегидрогеназа
ЛКП — локусы количественных признаков
МДА — малоновый диальдегид
МДГ — малатдегидрогеназа
МТ — металлотионеины
НиР — нитритредуктаза
НиРа — активность нитритредуктазы
HP — нитратредуктаза
ОСР — относительная скорость роста
ПВК — пировиноградная кислота
ПДК — пируватдекарбоксилаза
ПКС — протеинкиназная система
ПМ — плазмалемма
ПОЛ — перекисное окисление липидов
ПТ — продуктивность транспирации
ПЦР — полимеразная цепная реакция
ПФП — пентозофосфатный путь
ПЭГ — полиэтиленгликоль
РБФК — рибулозобисфосфаткарбоксилаза, или рибулозодифос-фаткарбоксилаза
РЦ — реакционный центр
СВЧ — реакция сверхчувствительности
СЖК — свободные жирные кислоты
СОД — супероксиддисмутаза
СС — сахарозосинтаза
ССК — светособирающий комплекс
СПУ — системная приобретенная устойчивость
ТКМ — термостабильность клеточных мембран
ТШ — тепловой шок
ТГП — температурный градиент посева
УДК — удельная длина корней
УДФГ — уридиндифосфатглюкоза
УКП — углекислотный компенсационный пункт
УМК — удельная масса корней
УПП — удельная поверхностная плотность
УППN — удельная поверхностная плотность азота
ФАР — фотосинтетически активная радиация
ФДК — фруктозодикиназа
ФП — фотосинтетический потенциал
ФС I — фотосистема I
ФС II — фотосистема II
ФХ — фитохелатины
ФЭИА — фотосинтетическая эффективность использования азота
цАМФ — циклический аденозин-монофосфат
ЦВ — ценотическое взаимодействие
цГМФ — циклический гуанозин-монофосфат
ЦТК — цикл трикарбоновых кислот
ЧПФ — чистая продуктивность фотосинтеза
ЩУК — щавелево-уксусная кислота
ЭИА — эффективность использования азота
ЭИВ — эффективность использования воды
ЭПА — эффективность поглощения азота
ЭПР — эндоплазматический ретикулум
ЭТЦ — электронтранспортная цепь
ABRE — элементы ответа на АБК
CDPK — Са-зависимая протеинкиназа
DRE — элементы ответа на дегидратацию
DREB 1 и DREB 2 — независимые семейства белков, связывающих элементы ответа на дегидратацию при низкой температуре (1) и засухе или засолении (2)
G-белки — ГТФ-зависимые белки
GPCR (G-Protein Coupled Receptors) — рецепторы, связанные с ГТФ-зависимыми белками
gs — устьичная проводимость
LEA (Late Embriogenesis Abundant) — белки позднего эмбриогенеза
МАРК — митогенактивируемая протеинкиназа
МАРКК — протеинкиназа, фосфорилирующая МАРК
МАРККК — протеинкиназа, фосфорилирующая МАРКК
Ppm (parts per million) — единица измерения относительных величин, равная миллионной доле
Ppb (parts per billion) — единица измерения относительных величин, равная миллиардной доле
PR-белки — белки патогенеза
PSS — фосфатидилсеринсинтаза
SOS (Salt OverSensitive) мутанты — сверхчувствительные к засолению мутанты
ВВЕДЕНИЕ
Растениеводческая отрасль страны функционирует в условиях изменения погодно-климатических факторов, возрастающей водной и ветровой эрозии почв при постоянном снижении биологического разнообразия в агроэкосистемах. При этом на эродированных землях водоудерживающая способность каждого гектара уменьшается на 500–600 м3, что равноценно снижению потенциальной урожайности зерновых культур на 5–12 ц/га. Вместе с тем для производства продуктов питания на 2500 ккал, которые ежедневно необходимы для питания только одного человека, уже требуется более 900 л воды. Кроме того, при существующих технологиях теряется, загрязняя окружающую среду, около 50–60% азотных, 70–80% фосфорных и свыше 50% калийных удобрений, до 60–90% поливной воды.
Многие факторы практикуемой в настоящее время системы интенсификации растениеводства, включающей использование высоких доз удобрений и пестицидов, видовую однотипность, генетическую однородность и загущение посевов, орошение и др., обычно снижают устойчивость агроэкосистем к действию абиотических и биотических стрессоров. Широкое применение пестицидов нарушает экологическое равновесие в агроэкосистемах (эффект «пестицидного бумеранга») и в большинстве случаев приводит к появлению более агрессивных и вирулентных рас патогенов, а также усилению вредоносности отдельных видов насекомых и сорных растений. Кроме того, техногенная интенсификация и узкая специализация хозяйств сопровождаются разрушением естественных элементов ландшафта, снижением разнообразия природных биотопов, уничтожением механизмов и структур биоценотической саморегуляции агроэкосистем.
Одновременно происходят глобальные и локальные изменения климата, и на территории страны участились повторяемость и продолжительность засух, в результате чего возросло число неурожайных лет. В XX столетии сильные засухи 14 раз поражали европейскую часть России и 8 раз – регионы Западной Сибири. В южных регионах страны летние засухи наступают каждый второй год с вероятностью 98%. В целом же в зависимости от погодных условий урожайность сельскохозяйственных культур изменяется в 2–3 раза в зонах устойчивого и в 5−6 раз в зонах неустойчивого увлажнения. В этих условиях необходимо уделять особое внимание подбору и размещению культур и сортов, применению техногенных средств, конструированию агроэкосистем и агроландшафтов. Игнорирование этих факторов снижает продуктивность посевов и, соответственно, эффективность производства. Действительно, если соотношение количеств энергии, запасенной культурами в урожае и затраченной на его формирование техногенной энергии, называемое коэффициентом энергетической эффективности (Кээ), составляло при экстенсивном хозяйствовании 20, то при современном интенсивном производстве полевых культур – 2, а в тепличном хозяйстве – 0,02.
Искомый коэффициент колебался для большинства зерновых и фуражных культур от 1,6 до 4,7, возрастая в следующем порядке: злаковые травы – сахарная свекла – кукуруза на силос – клеверозлаковая смесь – клевер красный. При этом даже при масштабном (тотальном) применении пестицидов потери урожая от вредных видов продолжают составлять 30–40%, снижая Кээ.
К настоящему времени земледелец располагает немалым арсеналом средств для увеличения потенциальной продуктивности растений (см. гл. 1 и 4). Действительно, урожайности сахарной свеклы в производственных посевах (США) и картофеля (Израиль) более 150 т/га, а также 14–18 т/га зерна кукурузы (США, Чили) уже не кажутся столь необычными. Однако при использовании агротехнических приемов, стимулирующих ростовые процессы, устойчивость растений к стрессорам, как правило, снижается. Так, в условиях даже орошаемого земледелия суховей в течение 2–3 ч, а стекание зерна в колосе непосредственно перед уборкой в течение недели могут привести к снижению урожайности на 50–90%.
Известно, что чем больше спектр и напряженность действия стрессоров (морозы, заморозки, засухи, суховеи, засоление, атмосферные загрязнители и др.) в той или иной агроэкологической зоне, тем ниже эффективность используемых удобрений и средств защиты растений. Поэтому конструирование агрофитоценозов и агроландшафтов, сочетающих высокую потенциальную продуктивность, экологическую устойчивость и средоулучшающие функции на основе целенаправленного формирования микрофитоклимата, зооценоза, сохранения механизмов и структур биоценотической саморегуляции, представляется весьма сложной задачей. Подобные системы должны включать многопольные севообороты, многовидовые агрофитоценозы, подпокровные, повторные и уплотненные посевы, подбор культур и сортов-взаимострахователей, а также поддержание экологического равновесия в агроэкосистемах на основе управления динамикой численности и изменчивостью структуры популяций полезных и вредных видов фауны и флоры и т. д. (Жученко, 2008). Рассмотрим вкратце, что представляет собой предлагаемая адаптивно-ландшафтная система.
Она, прежде всего, предполагает тщательный подбор культур и селекцию сортов, приспособленных для возделывания на бедных, кислых, уплотненных, засоленных и деградированных почвах с учетом агроэкологического районирования территории и конструирования агроландшафтов. Предлагаемая система, не отрицая важности применения минеральных удобрений, мелиорантов, пестицидов, регуляторов роста и пр., ориентирует на необходимость более полного использования ФАР путем биологизации и экологизации продукционного процесса в агроэкосистемах и агроландшафтах, мобилизации адаптивного потенциала агроценозов и эффективного использования всех ресурсов. Главные пути решения этой задачи связаны с расширением видового разнообразия агрофитоценозов, оптимизацией структуры посевных площадей и конструированием адаптивных агроэкосистем. Для этого необходим всесторонний учет отличающейся биоклиматической продуктивности различных сельскохозяйственных угодий и культивируемых видов (сортов) растений, средняя энергетическая продуктивность которых для разных территорий колеблется от 20 до 200 тыс. ккал на 1 м2 в год. В этой связи необходимо подчеркнуть преимущества смешанных, совместных, комбинированных, пожнивных, промежуточных, покровных и подпокровных посевов. Отметим, что КПД ФАР совместных посевов кукурузы и сои на 0,6% выше, чем в одновидовых посевах тех же компонентов.
Суммарная приходящая радиация в средних широтах за вегетационный период оценивается в 5 ‧ 109 ккал/га, из которой лишь около 1% используется на фотосинтез. Отношение формирующей урожай энергии ФАР (около 50% от приходящей) к потраченной техногенной (машинной) энергии составляет (2,5 ‧ 109 : 1,3 ‧ 106), или примерно 2000 : 1. Другими словами, на долю техногенной энергии в формировании урожая приходится лишь 0,05% от общих энергозатрат, что подтверждает непропорциональность прибавок урожая с увеличением затрат исчерпаемых ресурсов энергии.
Действительно, только одна или узкая видовая группа растений не могут эффективно использовать ФАР в течение всей вегетации. В этом, собственно, и состоит преимущество многовидовой структуры лугов и пастбищ (до 40–50 видов), обеспечивающих высокую фотосинтетическую продуктивность и экологическую устойчивость в течение всего вегетационного периода, в том числе и на начальных его этапах. Это особенно важно в условиях северных территорий России, где вегетация длится в течение всего лишь 60–90 дней, что накладывает существенные ограничения на использование биоклиматического потенциала региона.
Характерно, что в этих условиях ведущее место среди культурных растений по количеству накапливаемой за вегетацию сухой массы и энергии занимают лен (6540 млн ккал/га) и люпин (5134 млн ккал/га), наиболее приспособленные к почвенно-климатическим и погодным условиям Нечерноземной зоны. Особую ценность на этих территориях представляют также многолетние травы и насаждения, способные начинать утилизацию солнечной энергии сразу же после наступления температурного биологического минимума, а также позволяющие избегать вымывания питательных веществ при ежегодной вспашке и посеве однолетних культур.
Отметим, что различные виды растений и типы агроэкосистем характеризуются и разной степенью изменчивости величины и качества урожая в варьирующих условиях внешней среды. Так, если вариабельность урожайности многолетних трав в Нечерноземной зоне составляет в среднем 11% (максимум 20%), то у кукурузы и кормовой свеклы она в 2–2,5 раза выше. Валовые сборы зерновых культур в России также подвержены значительным колебаниям, размах которых (отношение максимального к минимальному сбору) варьирует в пределах 1,20−1,85. Этот же показатель за последние 15 лет составил в Германии − 1,05, Канаде и Мексике − 1,18, Австралии − 1.
Фитоценотическая совместимость разных культур и сортов, обусловленная спецификой их эволюционной и онтогенетической «памяти», а также необходимость синхронизации максимальной фотосинтетической продуктивности каждого агрофитоценоза с наиболее благоприятными для него местными условиями внешней среды, оказываются главным условием эффективного агроэкологического районирования. При этом особенно велика фитосанитарная роль севооборотов, которая существенно зависит от подбора предшественников, а также уровня плодородия почвы, и снижается при уменьшении содержании органических веществ в почве. Поэтому чем ниже плодородие почвы (кислые, засоленные, солонцеватые и др.), чем хуже климатические и погодные условия, тем меньше возможностей обеспечить благополучное фитосанитарное состояние агроэкосистем только за счет механизмов и структур биоценотической саморегуляции.
Использование в севооборотах не менее 3–4 сортов или гибридов рапса, овса, люпина, многолетних бобовых трав, различающихся по иммуногенетическим свойствам, позволяет улучшить фитосанитарное состояние посевов, в частности, значительно снизить поражение зерновых колосовых культур корневыми фузариозными гнилями. Корневые выделения ряда культур оказываются токсичными для нематод, проволочников и других вредителей. При возделывании многолетних растений для борьбы с фитонематодами используется естественная супрессивность почвы, обеспечивающая развитие полезной (ингибирующей, конкурентной, паразитной) микрофлоры.
В основе адаптивной стратегии развития сельскохозяйственного производства находится принцип предотвращения негативного эффекта стрессоров на посевы, как, например, предупреждение эпифитотий и эпизоотий на основе регуляции численности популяций полезной и вредной фауны и флоры в агроландшафтах. Соответствующие мероприятия, включающие диагностику, контроль, прогноз, сигнализацию и управление (регуляцию) указанной динамикой при их своевременном и качественном проведении, оказались наиболее эффективными. K примеру, мероприятия по предотвращению пагубных последствий засухи включают, наряду с районированием посевов и посадок, подбор засухоустойчивых культур и сортов (гибридов), сохранение запасов влаги за счет паров, мульчирование полей, использование кулис и лесополос, строительства ирригационных сооружений и т. д.
Считается, что за прошедшие 100 лет средняя температура на планете увеличилась на 0,6 °С. При этом высокие широты нагреваются больше, чем низкие, а сам нагрев выше зимой, чем летом. При общем росте количества осадков в теплом климате уменьшается число дней с осадками, но становятся заметны более продолжительные периоды интенсивных осадков, т. е. ливней. Одновременно возрастает и число дней без осадков. В результате повышается вероятность проявления как засух и длительных периодов жары, так и наводнений (Кошкин, Андреева, Гусейнов, 2019).
Хотя потепление на 1 °С и продвигает экономически оправданную границу возделывания зерновых культур в России на 300 км в более высокие широты (осеверение) и, возможно, станет позитивным фактором для умеренных зон, это не сможет компенсировать потерь урожая в основных зерновых районах, где усилится аридизация. Повышение же засухоустойчивости культур селекционными методами сопряжено с большими трудностями, что обусловлено решающим вкладом водного статуса растений в фотосинтетическую деятельность агрофитоценозов. Неслучайно абсолютное большинство засухоустойчивых видов растений отличается сравнительно низкой продуктивностью, а поиск соответствующих генетических доноров идет в направлении совершенствования архитектоники растений, усиления мощности корневой системы, опушенности листьев и стеблей и др.
Изменения отмеченных выше условий функционирования растениеводческого сектора, в том числе климата, предполагают внесение необходимых корректив в зональные системы ведения сельского хозяйства. Так, рост теплообеспеченности в Ставропольском крае сопровождался увеличением количества осадков, который в среднем по краю повысился за 50 лет на 80 мм, с колебанием от 31,4 до 130,1 мм. Большее количество осадков за период посев – всходы озимой пшеницы (на 33%) и весенне-летней вегетации (на 7–27%) – способствует росту урожайности этой культуры. Стал более жарким (+0,8 °С) и сухим (−3,9 мм) период созревания и уборки озимой пшеницы (июль), что способствует своевременной уборке и получению зерна высокого качества. Засушливость же осеннего периода требует выбора адекватных предшественников и перестройки системы подготовки почвы под посев озимой пшеницы.
Таким образом, всесторонний анализ и учет факторов, влияющих на рост, развитие и формирование урожая культур может содействовать созданию условий для более полной реализации потенциальной продуктивности и стресс-толерантности генотипов в достаточно широком диапазоне действия биотических и абиотических стрессоров.
Глава 1. ПОТЕНЦИАЛЬНАЯ УРОЖАЙНОСТЬ КУЛЬТУР ПРИ ОПТИМИЗАЦИИ ВЫРАЩИВАНИЯ И ПЕРЕДАЧА СИГНАЛОВ О ДЕЙСТВИИ СТРЕССОРОВ
1.1. Ретроспективный и перспективный анализ особенностей формирования урожая (на примере отдельных зерновых и зернобобовых культур)
Потенциальная урожайность достигается путем снятия лимитирования хода физиологических процессов факторами биотической и абиотической природы и использования всех экономически и экологически целесообразных техногенных средств интенсификации продукционного процесса. В связи с этим важно проанализировать, какие физиологические подходы можно использовать для достижения потенциальной урожайности. В приводимых ниже данных для сравнения использованы культуры с С3- и С4-типом фотосинтетического метаболизма углерода.
Кукуруза. Средняя урожайность кукурузы на зерно в штате Айова на площади 5,3 млн га составила 10,5 т/га, в том числе 0,5–0,7 т/га за счет трансгенеза, увеличиваясь начиная с 1996 года ежегодно на 214 кг/га, или 2% от данной урожайности, а потенциальная урожайность (ПУ) – на 1% (рис. 1.1). Что касается ПУ, она составила в США при орошении и густоте посева 100 000 раст./га, дозах азота 225–300 кг/га 16 т/га (Grassini et al., 2009), а в Чили – 20 т/га (Campos et al., 2004). Таким образом, сохраняется огромный разрыв между потенциальной и реальной урожайностью.
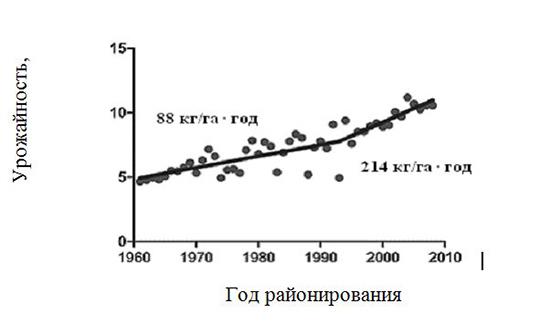
Рис. 1.1. Динамика урожайности кукурузы (т/га) в штате Айова (США)
В связи с вышесказанным интересны сравнения прибавки урожайности кукурузы и сои в штате Небраска за последние 40 лет прошлого столетия, составившие в среднем соответственно 98,0 ± 18,9 и 35,1 ± 5,3 кг/га ‧ год. Таким образом, урожайность кукурузы росла почти в 2,8 раза быстрее, чем сои. Примерно подобное соотношение сохраняется также при сравнении урожайности этих культур. Это объясняется тем, что семена сои, в отличие от кукурузы, более энергоемки и содержат большую сумму жира и белка (580 г/кг у сои и 150 – кукурузы). Это требует больших затрат АТФ и, соответственно, дыхательного субстрата (ассимилятов) для их синтеза, что снижает нетто-фотосинтез посева и урожайность сои. Это оказывает наряду с С4-типом фотосинтеза решающее влияние на преимущество кукурузы по эффективности использования радиации (ЭИР).
Определяющими в повышении урожайности современных гибридов является целый ряд факторов, в том числе более ранний посев, примерно на 12 дней раньше, чем в 1979 году, что позволяет посевам поглощать больше ФАР для налива зерна при более благоприятном температурном режиме и уходить от засухи в фазе цветения (см. гл. 3). Это достигнуто благодаря выведению холодоустойчивых, а также устойчивых к гербицидам и вредителям (болезням) гибридов. Кроме того, густота посева в кукурузном поясе США увеличивалась за последние 25 лет со скоростью 1000 растений/га ‧ год, достигнув в штате Айова 80000 раст./га за счет выведения гибридов, устойчивых к загущению, и эта тенденция сохраняется, несмотря на высокую стоимость семян (Kucharik, 2008). Активное использование технологий точного высева семян обеспечила выравненность проростков, а затем и взрослых растений (Edmeades et al., 2006). Вклад селекции в урожайность возрастает, и она уже превышает в последние годы 50%. Уже в ближайшие годы ожидается заметное повышение вклада биотехнологии, что вкупе с технологиями точного земледелия (см. гл. 9) может дать существенное увеличение урожайности кукурузы (рис. 1.2).
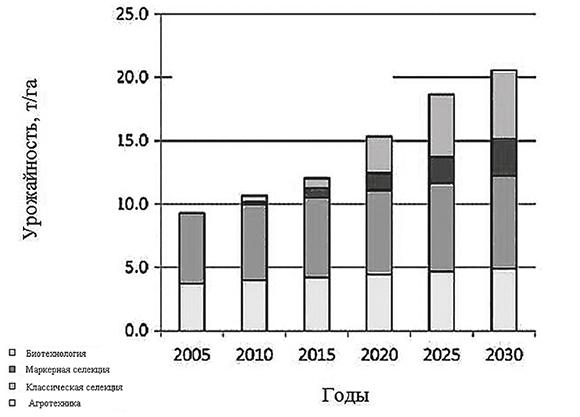
Рис. 1.2. Прогнозируемый вклад агротехники, традиционной селекции, маркерной селекции и биотехнологии в урожайность кукурузы в США (Edgerton, 2009)
В связи с вышеизложенным интересны данные по изменению параметров продуктивности гибридов кукурузы, созданных с 1930 по 2002 год (табл. 1.1). У последних гибридов отток ассимилятов в початок в критический период (одна неделя до выметывания нитей початка – 20 дней после этого) заметно выше, чем у гибридов ранней селекции. Именно с этого периода различия между ними по урожайности и КПД ФАР становятся заметнее. Кроме того, современные гибриды более устойчивы к ограниченной влагообеспеченности, даже во время цветения (см. рис. 3.21), а также к окислительному стрессу (Tollenaar and Yce, 2006).
Таблица 1.1
Изменения урожайности и морфофизиологических признаков гибридов кукурузы с 1930 по 2002 г. (Duwick, 2005)
| Показатель | Изменения показателя | Примечания |
| Урожайность при оптимальной густоте посева | Линейное увеличение | Около 80 кг/га в год |
| Оптимальная густота | > 80 000 раст./га | Началось с 30 000 раст./га |
| Биомасса | Постоянное повышение | |
| Кхоз | Небольшое повышение, сейчас 50–55% | Старые гибриды при высокой густоте отличаются пустозерницей |
| Число зерен на 1 м2 | Постепенное увеличение | Больше растений на 1 га, меньше пустозерницы |
| Масса 1 зерна | Небольшое увеличение | |
| Белковость зерна | Снижение | Увеличение крахмалистости зерна |
| Число дней до цветения |
Не изменилось | Число листьев постоянно |
| Период налива зерна | Длиннее | |
| Высыхание зерна в початке |
Быстрее | |
| Высота растений | Нет закономерного изменения | |
| Высота прикрепления початка | Небольшое снижение | |
| Полегание | Заметное снижение | Полностью не исключено |
| Посев | Более эректоидные листья | ИЛПмакс практически не изменился |
| Сохранение зеленых листьев при наливе | Значительно увеличилось | Небольшое изменение при терминальной засухе |
| Скручивание листьев* | Больше | Более очевидно у вертикальных листьев |
| Размер метелки | Сильно снизился | |
| Интервал между цветением метелки и выметыванием початка | Снизился почти до нуля | Индикатор стресс-толерантности при засухе |
* — cм. рис.1 из Приложения 1.
Что касается ИФ листьев, у старых и новых гибридов она существенно не различалась. Вместе с тем выравненность растений в посеве наряду с более эректоидными листьями обеспечивает более равномерное распределение ФАР по профилю посева и соответственно более стабильную ИФ у современных гибридов (Duvick, 1997).
Преимущество новых гибридов над старыми по урожайности составило 30% в выравненных и 49% – невыравненных посевах, увеличиваясь с повышением густоты и, соответственно, конкуренцией между растениями. При этом чем больше коэффициент вариации зерновой продуктивности отдельного растения в посеве, тем меньше урожайность (Tollenaar and Wu, 1999). Гибриды селекции 90-х годов прошлого века не отличаются по урожайности от сортов 30-х годов при выращивании при густоте посева 10 тыс. раст. / га (рис. 1.3), и в свою очередь урожайность последних не отличается при густоте 10 000 и 79 000 раст./га. (Duvick, 1997). При этом Кхоз составил около 50%, существенно не изменившись за последние 70 лет селекции, за исключением выращивания при густотах посева свыше 10 раст. /м2, когда он начинал снижаться. Из этого можно заключить, что урожайность зерна кукурузы, начиная с 1930 г., кратно возрастала главным образом за счет накопления большей биомассы и некоторого увеличения массы единичной зерновки на фоне примерного постоянства Кхоз (рис. 1.4).
Обращает также внимание повышение максимального значения индекса листовой поверхности (ИЛПмакс) с увеличением густоты посева. Однако данный фактор не оказывает решающего влияния на поглощение солнечной радиации. Так, повышение ИЛП с 3 до 4, т. е. на 33%, увеличивает поглощение лишь на 8%. Более весомой может быть роль эректоидности листьев, особенно верхних, у современных гибридов, что повышает равномерность распределения радиации по профилю посева (рис. 1.5). Показано, что повышение эректоидности листьев увеличивает урожайность на 20% (Tollenaar and Dwyer, 1997).
Что касается потенциальной ИФ в фазе выметывания нитей початка, она, как правило, выше у гибридов старой селекции, как и скорость ее снижения в фазе налива зерна (Ying et al., 2002). Это свидетельствует о том, что в благоприятных условиях выращивания ИФ не определяет преимущество современных гибридов по урожайности. Вместе с тем интенсивность текущего фотосинтеза листьев современных гибридов более устойчива к пониженным ночным температурам, недостатку влаги и азота, причем в обоих последних случаях снижение ИФ до нуля происходит раньше у ранних, чем у последних гибридов (Nissanka et al., 1997).
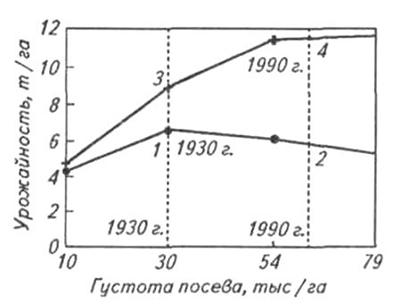
Рис. 1.3. Зависимость урожайности гибридов кукурузы 30–90-х годов XX в. от густоты посева (Evans and Fischer, 1999)
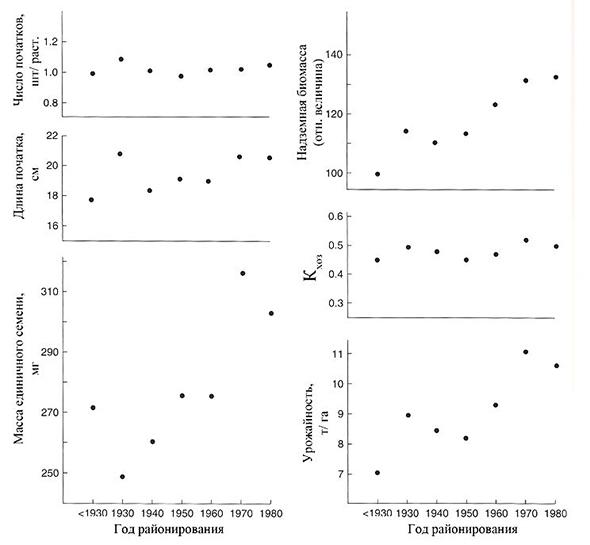
Рис. 1.4. Урожайность зерна и структура урожая сортов (гибридов) кукурузы, районированных в США в 1930-1980 гг. (до 1930 г. высевались сорта)
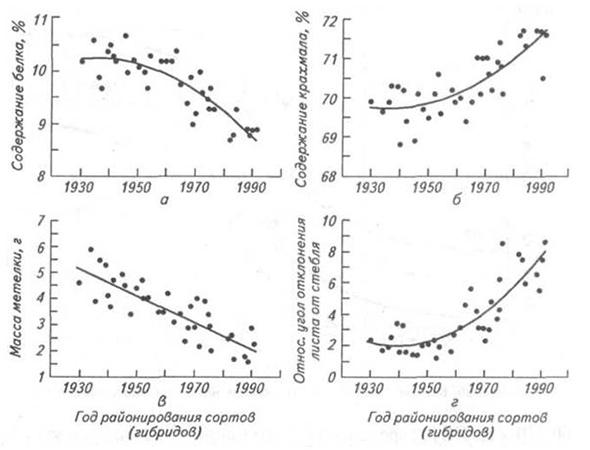
Рис. 1.5. Изменения содержания белка (а), крахмала (б) в зерне, массы метелки (в) и эректоидности листьев (г) во временной серии гибридов кукурузы. (Данные полевых опытов, проведенных в Айове с 1991 по 1994 г.; Duvick and Cassman, 1999)
Ориентация листьев кукурузы в загущенных посевах определяется реакцией на соотношение потоков красного и инфракрасного лучей (К/ИК) и направлена на минимизацию взаимного затенения листьев в посеве (Maddonni et al., 2002). Соотношение К/ИК в междурядьях у растений кукурузы выше, нежели в рядке. Интересно, что листья гибрида DК696 изменяют свою ориентацию в ответ на снижение соотношения К/ИК, а другого гибрида Exp 980 – не изменяют, т. е. наблюдается сортоспецифическая реакция. Опосредованными фитохромом реакциями на низкое соотношение К/ИК можно манипулировать для повышения урожайности, что установлено, в частности, для картофеля (Boccalandro et al., 2003). Экспрессия гена арабидопсиса PHYB, связанного с фитохромом В - фоторецептором, участвующим в детекции соотношения К/ИК в посевах разной густоты, повысила урожайность клубней трансгенного картофеля в поле. Таким образом, указанную манипуляцию можно использовать для создания сортов, устойчивых к выращиванию в загущенных посевах и соответственно более теневыносливых.
Пшеница. Ежегодная прибавка урожайности в мире за последние 20 лет составила 25 кг/га, что намного меньше, чем у риса – 38 и кукурузы – 80 кг/га (Fischer and Edmeades, 2010) (рис. 1.6). Вклад ежегодного повышения концентрации СO2 в атмосфере на 2 мкМ/моль (см.гл.13) в указанные прибавки может составить около 0,3%, по меньшей мере у С3-видов (пшеница и рис) (Tubiello et al., 2007).
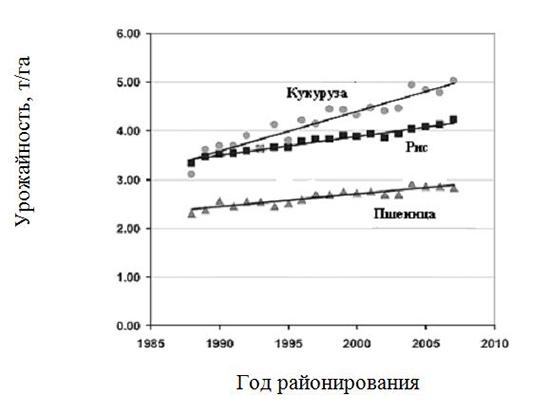
Рис. 1.6. Динамика среднемировой урожайности пшеницы, кукурузы и риса с 1988 по 2007 г. (ФАО, 2009)
В Великобритании в 2007 году средняя урожайность пшеницы при 15% влажности зерна на площади 2 млн. га составила свыше 8 т/га при внесении 190 кг/га N, 31 кг/га Р2О5 и 39 кг/га К20. С 1989 года урожайность росла со скоростью 53 кг/га в год, что составляет 0,7% от текущей урожайности. Полагают,что прогресс реальной урожайности на 90% обусловлен селекционным улучшением сортов (Маскау, 2009). При оптимальных условиях выращивания и использовании средств защиты растений урожайность в опытах в среднем с 2004 по 2008 год составляла 10,4 т/га, что может рассматриваться как потенциальная урожайность (ПУ). Примерно такая же урожайность достигнута при выращивании риса в производственных условиях в Японии, причем сорт Таканари (Takanari) был получен методом классической селекции. Реальная урожайность (РУ) пшеницы оказалась на 30% ниже ПУ (рис. 1.7). Заметим, что средняя урожайность пшеницы по России на площади более 27,5 млн. га составила в 2017 году около 3,0 т/га.
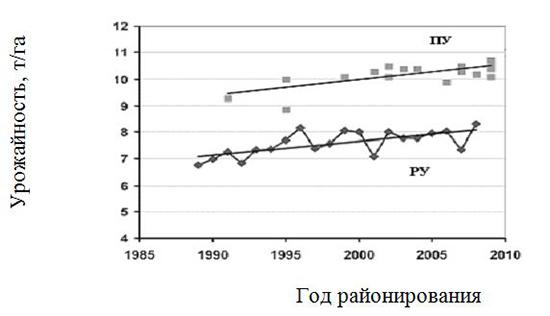
Рис. 1.7. Динамика потенциальной (ПУ) и реальной (РУ) урожайности пшеницы, выращиваемой в Великобритании (ФАО, 2009)
Увеличение урожайности пшеницы в процессе селекции связано скорее с изменением перераспределения надземной биомассы в зерно, чем с увеличением накопления надземной массы. При этом высота растения обычно отрицательно коррелирует с Кхоз. Однако есть пороговый уровень низкорослости растения, ниже которого урожайность за счет Кхоз далее уже не повышается. Поэтому для дальнейшего ее повышения необходимо вскрыть физиолого-генетическую основу повышения КПД ФАР и соответственно накопления надземной биомассы. Заметим, что современные сорта характеризуются более высоким КПД ФАР, чем их предшественники, во вторую половину вегетации, сопровождаемым увеличением биомассы после цветения. Это может быть, в частности, следствием повышенного спроса на ассимиляты у сортов с более крупными колосьями в сравнении с сортами ранней селекции, что повышает ИФ у современных сортов (Reynolds et al., 2004). Это обусловлено большей устьичной проводимостью для молекул углекислоты.
Признаки, улучшающие стеблестой. Крупнозерные генотипы пшеницы обладают большей всхожестью и более крупными проростками, чем мелкозерные (табл. 1.2 и подробнее в табл. 3.4, гл. 3). Однако норма высева обычно рассчитывается в ц/га, поэтому крупнозерность может и не дать очевидных преимуществ. Самыми эффективными являются отбор на ширину листьев колеоптиле и размер колеоптильных побегов, которые должны отбираться вместе с длиной колеоптиле первыми. В этой таблице наряду с главными (высокой приоритетности) показателями приведены вторичные (менее важные) показатели. Хотя генетическая вариация по последним также существует, она незначительна и по ним трудно вести отбор. Тем не менее, экспрессия этих признаков может быть в некоторых случаях улучшена в процессе селекции на первичные признаки. Так, например, для улучшения стеблестоя путем увеличения длины колеоптиле рекомендуется замена нечувствительных к ГК генов карликовости Rht1 и Rht2 чувствительными к ГК генами. Результаты тестов показывают, что это увеличивает также всхожесть и скорость разворачивания листьев, что означает начало их активного фотосинтеза. Более того, селекция на широкие первичные (зародышевые) листья, интегрирующие как размер зародыша, так и УПП листа, должна также повышать относительную площадь листьев, рассчитываемую как отношение площади листьев к сухой надземной массе побега (дм2/г). Важно, что гены карликовости, чувствительные к ГК, хорошо наследуются и по ним хорошо вести отбор.
Таблица 1.2
Признаки, потенциально улучшающие стеблестой и раннее его формирование у пшеницы (Richards et al., 2001)
| Признаки | Насле- дуемость |
Ожидаемый эффект взаимо- действия G x Е |
Универ- сальный/ средоза- висимый признак |
| Главные | |||
| Длинные колеоптили | высокая | универ- сальный |
|
| Широкие листья проростков | высокая | средо- зависимый |
|
| Размер зародыша | высокая | средо- зависимый |
|
| 1/УПП | средняя | ||
| Крупный колеоптильный побег | средняя | средо- зависимый |
|
| Вторичные | |||
| Крупные зерна | высокая | универ- сальный |
|
| Быстрое появление всходов | низкая | средо- зависимый |
|
| Высокая скорость разворачивания листа | средняя | средо- зависимый |
|
| Холодоустойчивость | средняя | средо- зависимый |
|
| Глубина заложения узла кущения | средняя | средо- зависимый |
|
| Относительная площадь листьев | средняя | средо- зависимый |
Хорошая наследуемость длины колеоптиле показывает, что скрининг этого признака можно начинать уже в F2 поколении.
Сравнение силы роста пшеницы и ячменя показало, что площадь листьев проростка у ячменя почти в 2 раза больше, чем у пшеницы, прежде всего за счет более раннего прорастания (на 1 сутки), более крупного зародыша и меньшей толщины листьев (1/УПП). Изучение мировой коллекции пшениц обнаружило две популяции, показывающие аналогичные с ячменем данные на уровне проростков (Richards and Lucas, 1991). Cовмещение двух независимых признаков – большего зародыша и более высокой относительной площади зародышевых листьев – дало потомство даже с большей силой роста, чем у родителей (Richards, 1996). Однако наследуемость признака раннего формирования большей площади листьев оказалась невысокой. Вместе с тем средняя ширина первых двух листьев хорошо наследуется и имеет тесную генетическую корреляцию с площадью листьев, что позволяет использовать первую как индикатор при селекции на увеличенную площадь листьев, тем более что ширина, в отличие от площади листьев, менее чувствительна к взаимодействию генотипа и среды.
Повышение КПД ФАР. Ежегодные темпы повышения урожайности, необходимые для ее удвоения, к середине столетия должны составить от 1,16 до 1,31% в среднем по всем зерновым культурам и 1,7% – по пшенице (Hall and Richards, 2013). Вместе с тем реальные темпы прироста за последние 20–30 лет намного скромнее и составляют, в частности, для риса и пшеницы всего около 1% (Ray et al., 2013), сои – 1,3% и кукурузы 2,6%. Вклад повышения температуры в искомые величины в результате глобального потепления оценивается примерно в 30% (Lobell and Field, 2007). В среднем за вегетационный период эффективность трансформации ФАР в биомассу у современных сортов пшеницы достигает 2,5 г сухой массы/МДж (для сравнения 2,2 для риса и 3,3 – кукурузы) (Bingham et al., 2007). Максимальное значение эффективности использования радиации (ЭИР) у кукурузы объясняется высокой интенсивностью С4-фотосинтеза, а минимальное у риса – высокой интенсивностью фотодыхания.
Теоретический максимум КПД ФАР, составляющий 0,094 (C3 виды) и 0,123 (C4 виды) намного превышает реальные величины, в связи с чем данный показатель представляется идеальным кандидатом для повышения потенциала урожайности (Zhu et al., 2010). Однако он варьирует в широком диапазоне и сильно зависит от факторов среды (изменений климата) и культур (Slattery et al., 2013). Одним из главных путей повышения КПД ФАР на уровне листа является увеличение эффективности карбоксилирования ключевым ферментом РБФК при одновременном снижении фотодыхания (Raines, 2011; Evans, 2013). На уровне посева решающими факторами являются архитектоника растений и размеры антенны светособирающего комплекса (CCК) для улучшения распределения квантов света в плотных (загущенных) посевах (Ort et al., 2011; Ainsworth et al., 2012), что оптимизирует распределение N по ярусам листьев, а также повышает ИФ колоса (Reynolds et al., 2012).
Кукуруза, обладающая высокой скоростью роста, демонстрирует самое высокое значение КПД ФАР среди кормовых культур, равное 0,0488, что составляет менее половины от теоретического максимума 0,123. Несколько меньшее значение установлено для другого растения с С4 типом фотосинтеза сорго – 0,0455, что составляет около 37% от максимального значения. Среди небобовых культур с C3-типом фотосинтеза наибольшее значение искомого показателя отмечено у картофеля – 0,0414. Бобовые культуры, напротив, характеризовались минимальным среди C3 видов значением 0,028, что составляло 31% от возможного максимума для C3 видов. При учете подземной массы у однолетних культур значение коэффициента повышалось на 10–20%. Однако даже в этом случае у картофеля он составил лишь 44% от теоретического максимума. У сои энергия, заключенная в биомассе корней и клубеньков, достоверно выше, чем у небобовых культур, что повышает КПД, но в лучшем случае до 0,03. Последнее значение тем не менее ниже такового для небобовых культур с C3-типом фотосинтеза. Это можно объяснить затратой энергии на симбиотическую азотфиксацию (Andrews et al., 2009), на которую уходит от 9 до 17% ассимилированного углерода (Fujikake et al., 2003; Ito et al., 2006). Если принять это во внимание, КПД может увеличиться до 0,034, что все еще ниже, чем у небобовых культур. Таким образом, имеются какие-то иные факторы, его ограничивающие у зернобобовых культур. К таковым можно отнести, например, запаздывание с формированием зрелого симбиотического аппарата (клубеньков) в начале вегетационного периода. В связи со сказанным, повышение эффективности симбиотической азотфиксации может стать важным инструментом увеличения КПД. Вместе с тем он у земляного ореха – представителя бобовых культур – намного выше, чем, например, у сои, и равняется таковому у небобовых культур. Подобная аномалия объясняется тем, что энергоемкость растения арахиса в 1,2 раза выше, чем у других бобовых культур (Slattery and Ort, 2015).
Заслуживает также внимания значение КПД ФАР у нового гибрида риса (0,0472) и сортов индийского подвида риса (0,0442), что заметно выше, чем у сортов японского подвида (0,0388) и аналогично данным у кукурузы. Анализ показывает, что подобные высокие значения КПД ФАР у нового гибрида и индийского подвида риса объясняются не генетическими особенностями, а условиями выращивания. Озимая пшеница (0,0399) также заметно превосходит яровую (0,0352).
Необходимо заметить при этом, что новые гибриды риса отличаются ограниченным кущением, в результате чего даже в загущенных посевах свет хорошо проникает к расположенным в нижних ярусах листьям (Peng et al., 2008). Изменение концентрации пигментов или архитектоники посева при высоком значении ИЛП может оптимизировать доступность квантов света для листьев разных ярусов, снижая таким образом неэффективное использование приходящей солнечной радиации.
Энзим РБФК/о, катализирующий ассимиляцию СО2 в цикле Кальвина, является наиболее очевидной мишенью для увеличения ИФ и, следовательно, КПД ФАР у С3 растений. Однако РБФК/о проявляет слабую каталитическую активность, поэтому для поддержания высокой ИФ требуются значительные его количества. Данный фермент участвует также в окислении рибулозобифосфата, инициируя фотодыхание и последующее выделение СО2, NH3, а также повышая расходную часть энергетического баланса клетки. Подобная двоякая роль РБФК/о сильно влияет на эффективность транспирации. Это обусловлено тем, что закрытие устьиц при ограниченной влагообеспеченности приводит к понижению соотношения концентраций СО2/О2 в клетке, делая ассимиляцию углерода менее эффективной, и наоборот, полностью открытые устьица будут усиливать потери воды. Поэтому высокие температуры и даже засуха (или их сочетание) могут в дальнейшем увеличивать окислительную активность РБФК/о относительно карбоксилирования РБФ и, соответственно, усилить фотодыхание (Parry et al., 2007).
За исключением кукурузы и сорго все основные культуры, такие как пшеница, рис, ячмень или соя, имеют С3 тип фотосинтеза. Существует несколько путей увеличения ассимиляции СО2 и, соответственно, снижения потерь при фотодыхании. В то время как один из подходов предполагает внедрение форм фермента с высоким сродством к СО2 в С3 растения, альтернативные направления предполагают перенос механизма концентрирования СО2 по типу С4 фотосинтеза в С3 растения. Подобный подход был бы особенно выгоден при засухе или в условиях, ведущих к закрытию устьиц и соответственно к снижению внутриклеточной концентрации СО2 (von Caemmerer, 2003). Иные подходы по увеличению ИФ влияют на регенерационную способность РБФ и включают в себя другие ферменты цикла Кальвина, и особенно седогептулозо-бифосфатазу а также компоненты электронного транспорта и светособирающего комплекса (Parry et al., 2007, 2008). Увеличение карбоксилазной активности по отношению к окислительной путем изучения мутантных или химерных ферментов РБФ даже на бактериях или на водорослях пока что не было достигнуто. Вместе с тем, на модели показано, что замещение РБФК/о у пшеницы на РБФК/о от Limonium gibertii может увеличить фотосинтез на 12% и повысит урожайность. Несмотря на большой прогресс в разработке методов перенесения генов у элитных сортов пшеницы, успех в трансформации пластидного генома, поскольку указанный фермент находится в хлоропластах, весьма скромный и ограничивается пока лишь модельными объектами.
Другим альтернативным подходом для преодоления ограничений РБФК/о при С3 типе фотосинтеза может быть концентрирование СО2 в каталитическом центре. На рисе были сделаны некоторые попытки в этом направлении, однако сложность анатомии листа С4 растений и кооперативный фотосинтез с участием клеток мезофилла и обкладки проводящих пучков препятствует перенесению подобной системы в С3 растения. Открытие СО2 насоса и возможности манипуляции им в отдельных клетках привели к разработке ряда биотехнологических методов по перенесению насосов отдельных клеток в С3 культуры, такие как рис и пшеница (Leegood, 2002). Однако данный подход все еще требует более детального изучения координированной экспрессии многих трансгенов, сборки генпродуктов в корректных зонах на клеточном и субклеточном уровне при соответствующей регуляции активности ферментов и транспортеров. Дополнительная сложность обусловлена со сборкой копий большой и малой субъединицы РБФК/о в функциональный энзим in vivo, что требует высокого уровня экспрессии, взаимодействия с шаперонами, посттрансляционной модификации и взаимодействия с активазой РБФК/о (Parry et al., 2007).
Биотехнологические подходы были использованы так же для перенесения ферментов бактерий в фотодыхательный цикл растений с целью уменьшения тем самым выделения NH3 (Parry et al., 2003). Метаболический путь превращения хлоропластных гликолатов непосредственно в глицерат без участия NH3 был внедрен в арабидопсис, причем двуокись углерода выделялась непосредственно в хлоропласт. Это сопровождалось увеличением фотосинтеза, в том числе за счет повышения концентрации СО2 в активном центре РБФК/о (Kebeish et al., 2007).
Другие методы включают предотвращение снижения активности РБФК/о при высокой температуре путем увеличения концентрации и термостабильности активазы данного фермента, а также предотвращения его деградации или содействия ее синтезу (Parry et al., 2008)
Фосфоенолпируваткарбоксилаза (ФЭПК), которая служит в качестве помпы, концентрирующей CO2 при С4 фотосинтезе, специфично экспрессируется в клетках мезофилла листьев. Регуляция экспрессии данного фермента в клетках мезофилла С3 видов представляет собой еще одну альтернативную мишень для увеличения ИФ последних.
Накопление биомассы. Небольшое увеличение надземной биомассы отмечено у отдельных последних сортов яровой и озимой мягкой пшеницы в сравнении с сортами ранней селекции (Shearman et al., 2005). Это обычно связано с некоторым повышением ИФ в период роста колоса и более значительным – при наливе зерна. Тот факт, что число зерен в колосе можно искусственно повысить путем короткого воздействия на растения светом высокой интенсивности в период быстрого роста колоса, указывает на то, что потенциальная фотосинтетическая способность элитных линий превышает уровень, необходимый для нормальной закладки и налива зерна. На кукурузе увеличение потенциальной урожайности сопровождалось увеличением общей биомассы на фоне относительно стабильного значения Кхоз. Большее накопление сухой массы новыми гибридами по сравнению с ранними можно объяснить более продолжительной фазой налива зерна у первых при примерно одновременном наступлении цветения початка и созревания у ранних и новых гибридов (см. гл. 3).
Фенологическое регулирование. Альтернативным путем увеличения числа зерен на единицу площади посева может быть удлинение фазы элонгации стебля. Предполагается, что это приведет к накоплению большей сухой массы колоса и, соответственно, к увеличению числа налившихся зерен. Однако это положение верно только в том случае, если продолжительность этой фазы можно регулировать и если ожидаемый прирост биомассы в течение этого периода не компенсируется уменьшением Кхоз. Подобные манипуляции могут включать гены, ответственные, в частности, за чувствительность к фотопериоду или раннеспелость. Гипотетическое участие генов фотопериода подтверждается тем фактом, что продолжительность элонгации стебля регулируется фотопериодической реакцией. Механизм, с помощью которого фотопериод изменяет количество зерновок на 1 м2, является таким же, каким влияет на него количество поглощенной радиации в фазе элонгации стебля (Gonzalez et al., 2005). Таким образом, идентификация и последующие манипуляции с генами, детерминирующими чувствительность к фотопериоду в фазе элонгации стебля, могут быть в будущем эффективным подходом для увеличения потенциальной урожайности.
Метаболизм азота. Удвоение производства сельскохозяйственной продукции в мире достигнуто за счет семикратного увеличения внесения азотных удобрений (см.гл.9). Однако из-за экологических и экономических причин дальнейшее расширение производства продуктов питания должно обеспечиваться благодаря более эффективному использованию азотных удобрений. Последнее особенно актуально для злаков, эффективность использования азотных удобрений под которые составляет около 35% (Hirel et al., 2007). Современные исследования на мягкой пшенице, кукурузе и рисе направлены на идентификацию ключевых признаков, определяющих формирование урожая на низком фоне азотных удобрений, выявление хромосомных участков и генов, вовлеченных в устойчивость к недостатку азота (Laperche et al., 2007). Так, методами количественной генетики установлено, что вариации в использовании азота гибридами кукурузы, включая его ремобилизацию, на низком фоне азота были больше, чем в поглощении азота до или после цветения, а на высоком фоне, напротив, меньше (Gallais and Coque, 2005).
Изучение влияния нокаут-мутаций на урожайность зерна и структуру урожая кукурузы и пшеницы при субоптимальных дозах N удобрений показало, что урожайность зерна контролируется активностью глутаминсинтетазы (Martin et al., 2006; Bernard et al., 2008). Ключевая роль этого фермента независимо от уровня азотного питания была определена путем идентификации хромосомного региона, ответственного за азотное питание.
Соя. Урожайность сои в CША с 1993 по 2006 год увеличилась с 2,2 до 2,9 т/га, а в других крупных странах-производителях она оказалась примерно на 0,7 т/га ниже. Максимальная урожайность сои в США на оптимальном агрофоне на площади 2 га составила 6,66 т/га, или 83% от предполагаемых теоретического максимума 8 т/га (Sinclair, 1999). Согласно модели, в производственных условиях средняя урожайность в 4 т/га, что на 60% выше средней урожайности 1998 года, может быть достигнута к 2043 году при широком использовании гибридных форм. Китайским селекционерам после 20 лет работы в 2003 году первыми в мире удалось создать гибридную сою.
При сравнении 16 сортов сои разных лет селекции показано, что сорта с высокой урожайностью отличаются также высоким значением Кхоз (Specht et al., 1986). В другом исследовании (Frederik et al., 1991) установлена зависимость Кхоз от года районирования сорта как в оптимальных условиях выращивания, так и при засухе. Вместе с тем прямая селекция на повышенный Кхоз при выведении новых сортов оказалась бесперспективной, хотя раннеспелые сорта, например, могут иметь более высокий Кхоз, но отличаются низкой урожайностью (Sinclair, 1993). Смена стратегии селекции от недетерминантных к детерминантным (или полудетерминантным) также не оказывает заметного эффекта на Кхоз (Frederik and Hesketh, 1994). Считается, что резерв дальнейшего увеличения Кхоз с целью повышения урожайности практически исчерпан и речь в будущем может идти главным образом о накоплении большей биомассы. Оно определяется количеством поглощенной радиации, эффективностью ее использования и/или длительностью вегетационного периода. Первый из указанных трех параметров определяется быстротой смыкания междурядий, густотой и временем посева. Показаны различия между сортами по быстроте начального роста и смыкания междурядий в посеве (Mian et al., 1998). Преимущество в энергии прорастания может обеспечить быстрое появление и разворачивание листовой пластинки, сокращая таким образом период, необходимый для формирования посева, с ИЛП=3,3. Это требуется для улавливания 90% падающей радиации.
Современные сорта отличались также более высокими содержанием хлорофилла и УПП листьев, хотя корреляция между данными показателями была несущественна. ИФ в фазе налива семян у современных сортов была также выше, чем у сортов ранней селекции (рис. 1.8), не отличаясь в фазе вегетативного развития. Урожайность последних ограничена мощностью донора при наливе семян из-за меньшего значения ФП. Действительно, при удалении части бобов у сортов разных лет селекции масса семян в оставшихся бобах у современного сорта увеличивалась намного больше, чем у сортов ранней селекции.
Горох. Генетический потенциал растений гороха и биоклиматические ресурсы Центральной России способны обеспечить формирование семян свыше 15 г/раст., или 12 т / га. Исходя из этого, селекционеры могут рассчитывать в отдаленной перспективе на создание зерновых сортов культуры с урожайностью 7...8 т/га при густоте стояния растений 90…100 шт./м2. В производственных опытах у районированных сортов уже сегодня достигнута урожайность 6,22...6,63 т/га, что близко к запланированному (Амелин и др., 2004).
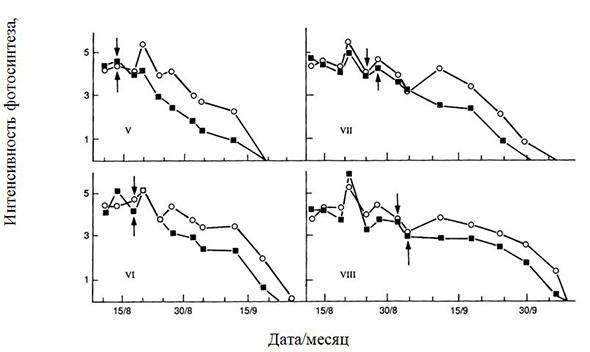
Рис. 1.8. Динамика интенсивности фотосинтеза (г СО2/м2 час) посевов у высоко (○) - и низкопродуктивных (■) сортов сои разной скороспелости (V – ультраскороспелый, VIII – исключительно позднеспелый). Стрелки указывают на начало налива бобов
Рост урожайности гороха в процессе селекции достигался главным образом за счет повышения Кхоз. при относительно стабильном уровне накопления сухой массы растениями в агрофитоценозе. Однако уборочный индекс у гороха тоже уже приблизился к максимально возможной величине, превышая у лучших сортов 0,55. Величина сухой фитомассы 12–15 г/раст. определена на период уборки из расчета, что при уборочном индексе 0,55...0,60 8–9 г/раст. будет приходиться на семена. В модельных полевых опытах сухая надземная масса составляла от 20 до 30 г/раст.
Вместе с тем в производственном агроценозе растения современных сортов накапливают в среднем 7...8, в лучшем случае – 9...10 г/раст., что составляет лишь 35–45% от возможного, и в результате длительной селекции эта ситуация фактически не улучшается. Таким образом, для создания сортов гороха с биомассой 12...15 г усилия селекционеров должны быть направлены, в первую очередь на повышение активности фотосинтетического аппарата и корневой системы растений.
Перспективным для этих целей может быть морфотип «хамелеон», растения которого формируют одновременно и листочки, и усики (рис. 1.9). Феномен гетерофиллии способствует накоплению большей по сравнению с сортами листочкового и усатого типа биомассы, соответственно на 10... 12 и 25...37%. Это сочетается со всеми основными хозяйственно-ценными признаками, что открывает возможность создания сортов, приближенных к идеатипу. Явление гетерофиллии обусловлено взаимодействием генов аf и tас, расположенных в одной хромосоме.
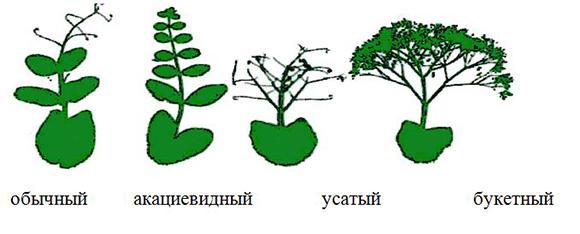
Рис.1.9. Морфотипы гороха
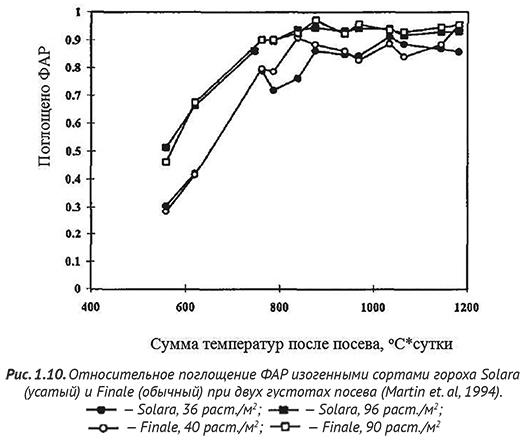
Низкорослые сорта гороха с короткими междоузлиями, цепляясь усиками образуют устойчивый к полеганию стеблестой, технологичные свойства которого определяются жесткостью и прочностью стебля, упругостью и строением усиков, размерами прилистников. Обладая меньшей фотосинтезирующей поверхностью, сорта с усатыми листьями имеют интенсивность фотосинтеза не меньше, чем обычные, так как не полегают, и благодаря этому стеблестой у них лучше освещен и лучше проветривается. Как показано на рис. 1.10, при контрастных густотах посева у обычной и усатой форм гороха не обнаружено заметной разницы в течение вегетационного периода по количеству поглощенной ФАР. Это свидетельствует о существенной роли усиков гороха в поглощении радиации и, соответственно, фотосинтезе целого растения. Однако при засухе и переувлажнении усатые формы резко снижают урожайность. При этом эффективность использования радиации (ЭИР) зависит от напряженности донорно-акцепторных отношений в онтогенезе растений (рис. 1.11), увеличиваясь по мере роста бобов и снижаясь с завершением налива семян (Jeuffroy and Ney, 1997).
Площадь и УПП листьев. В отличие от других сельскохозяйственных культур (пшеница, ячмень, картофель) в селекции зернового гороха наблюдается четко выраженная тенденция уменьшения размеров листовой поверхности у новых сортов за счет числа листьев, а в отдельных случаях, и за счет их размеров. При этом листовые пластинки и прилистники становятся значительно толще и плотнее (на 19...26%), а черешки нижних и верхних листьев – короче (в среднем на 20%), средних – длиннее (в среднем на 14%) при более остром угле отхождения от стебля в генеративной сфере, что может повысить КПД ФАР
Достигнутая в настоящее время у листочковых сортов гороха площадь листовой поверхности (400...600 см2/раст.) при густоте стояния растений 90...100 шт. на м2 является близкой к оптимальной. Однако уменьшение последней без снижения числа листьев и конечного урожая еще возможно путем создания сортов мелколисточковых или с «усатым» типом листа при наличии у них хорошо развитых прилистников.
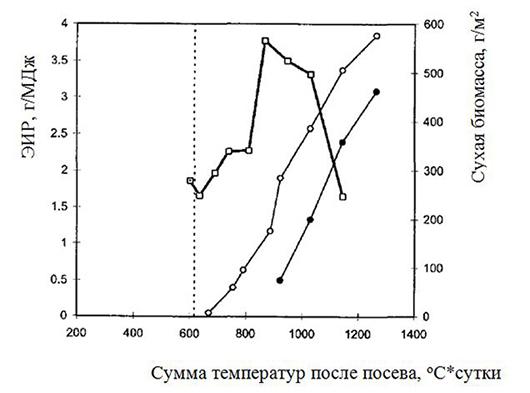
Рис. 1.11. Эффективность трансформации радиации в биомассу (ЭИР) и накопление биомассы бобов и семян посевами гороха усатого морфотипа (Martin et. al, 1994). 1 – ЭИР; 2 – сухая масса семян; 3 – сухая масса бобов; ...... - цветение
В селекции гороха, как и у многих других культур, площадь и УПП листьев коррелируют отрицательно (r = –0,98). При этом сортообразцы с более высокой УПП листочков и прилистников растений, как правило, отличаются повышенной семенной продуктивностью – r = +0,84...0,71 и интенсивностью фотосинтеза.
ЧПФ и ФВАХ. Урожайность зерна коррелирует с величиной ЧПФ, достигающей в среднем за вегетацию достаточно высокой величины 7–8 г/м2 сутки. Величины ЧПФ и фотовосстановительной активности хлоропластов (ФВАХ) за длительный период селекции зернового гороха возросли более чем на 30%, причем последняя у усатых форм оказалась выше, чем у обычных (соответственно около 14 и 10 моль К4Fе(СN)6/моль хл. мин).
Увеличение урожайности посевов гороха в будущем более чем на 50% связано с увеличением ЧПФ на 30–50% уже на ранних этапах развития, что должно обеспечить им формирование большего фонда ассимилятов в фазе вегетативного развития. Однако это представляет исключительно сложную задачу.
Ввиду того, что ЧПФ тесно связана с ФВАХ (r = +0,63), при создании сортов гороха, соответствующих идеатипу, актуальным является учет и последнего показателя, который генетически детерминирован и мало зависит от условий среды. Содержание хлорофилла в органах растений в данном случае не лимитирует как ИФ, так и продуктивность.
В селекции сортов усатого типа из-за небольших размеров листовой поверхности ФВАХ важно поддерживать на высоком уровне не только в листьях, но и в других фотосинтезирующих органах растений, в частности в усиках, особенно в период налива семян.
Параметры корневой системы и азотфиксация. Повышение урожайности в процессе селекции гороха сопровождалось увеличением мощности корневой системы на 20–30% на фоне уменьшения ФП вследствие интродукции в генотип рецессивных генов af и st. При этом сорта с усатым типом листа уступали листочковым по массе и объему корней в посеве на 10–28%, площади общей и поглощающей поверхности – 13–35%, длине – 16–40%. В целом выявлена тесная сопряженность (r = 0,78–0,98) размеров активной поглощающей поверхности корней и площади фотоассимилирующих листьев. Поэтому безлисточковые сорта уступают листочковым по мощности корневой системы даже при равной урожайности, а высокая потребность растений в элементах питания удовлетворяется за счет более интенсивного их поглощения (в 1,5–1,6 раза) в расчете на 1 м2 поглощающей поверхности корня.
Вместе с тем некоторые современные, хорошо окультуренные сортообразцы имеют высокие показатели как азотфиксирующей, так и производственной эффективности. Так, сорт Зубр сочетает высокую отзывчивость на инокуляцию семян Rhizobium (прибавка урожая зерна 7,8%) с высокой урожайностью (в среднем за 3 года 4,79 т/га). Этот пример подтверждает принципиальную возможность проведения целенаправленной селекции на повышение эффективности симбиотической азотфиксации у культуры.
Морфотипы и условия выращивания. В регионах с засушливым климатом и низкой культурой земледелия наиболее предпочтительны листочковые сорта с физиологически ограниченным ростом в период налива семян, тогда как в благоприятных погодных и агротехнических условиях произрастания безлисточковые сорта с генетически детерминированным типом развития стебля (3...5 фертильных узлов). При этом для селекции сортов усатого типа важно учитывать, что искомая детерминантность сопряжена с редукцией прилистников в зоне плодоношения растений и снижением их доли в биомассе ниже допустимых пределов – до 11–13%, против необходимых 18–22%.
Редукция прилистников в генеративной сфере при отсутствии настоящих листочков у сортов с усатым типом листа уменьшает биосинтетический потенциал продуктивного узла и растения в целом, особенно при неблагоприятных погодных условиях. Поэтому одна из возможностей совершенствования безлисточковых детерминантных генотипов заключается в придании способности формировать крупные парные прицветнички. Введение в генотип гороха гена brac, контролирующего этот признак, приводит к увеличению продуктивности усатых детерминантов на 33%.
В целом необходимо стремиться к созданию сорта с максимальным разделением вегетативной и генеративной фаз развития. После окончания цветения у них не должно происходить образование новых листьев и удлинение стебля с тем, чтобы все образующиеся ассимиляты транспортировались в семена.
У зернового гороха листочкового типа нежелательно иметь и сорта с продолжительностью вегетационного периода более 75 дней, поскольку высокая влажность почвы в конце июля – начале августа может усилить вегетативный рост и соответственно повысить опасность полегания.
У зерновых сортов усатого типа с детерминантным типом роста и повышенной устойчивостью к полеганию вегетационный период, напротив, может быть увеличен до 78 дней за счет удлинения репродуктивного развития, чтобы усилить семяобразование.
Интересно, что при сравнении двух больших групп – с обычными листьями и усатыми, в которых листочки видоизменены в усики, среднерослые сорта как с признаком неосыпаемости, так и без него давали примерно равный урожай зерна.
Устойчивость к полеганию. Основная причина раннего и сильного полегания посевов культуры – формирование у растений длинного и неплотного стебля с тонкими и длинными междоузлиями. Доказано, что для центральных областей России оптимальным является создание сортов с длиной стебля от 60 до 90 см. Формы с более коротким стеблем снижают урожайность, особенно в засушливые годы. Отбор форм с длиной стебля более 100 см также не рекомендуется, так как они характеризуются не только слабой устойчивостью к полеганию, но и низкой величиной Кхоз, составляющей 0,12...0,25.
По мнению А.В. Амелина с сотр. (2004), наиболее значимым морфофизиологическим показателем при отборе устойчивых к полеганию генотипов является линейная плотность стебля (ЛПС) или сухая масса единицы длины стебля (мг/см), независимо от морфотипа и целевого использования. Между линейной плотностью стебля и устойчивостью к полеганию у сортов гороха установлена достоверная положительная связь – r = +0,82. Не случайно в процессе селекции культуры с ростом этого показателя у растений отмечалось повышение как устойчивости к полеганию, так и продуктивности.
Современные зерновые сорта гороха характеризуются ЛПС во время уборки от 14 до 18 мг/см, что обеспечивает устойчивость к полеганию лишь до фазы зеленой спелости нижних бобов. Дальнейшее увеличение ЛПС на основе существующей тенденции в селекции этой культуры может помочь решить проблему полегаемости.
Увеличения урожая семян у будущих сортов гороха следует добиваться не за счет повышения их крупности, а посредством улучшения у растений плодо- и семяобразования. Действительно, в настоящее время масса 1000 семян составляет 280 г и выше. В результате укрупнения семян под действием искусственного отбора в них произошло существенное снижение относительного содержания белка, отдельных незаменимых аминокислот, зольных элементов при возросшем поражении их бактериозом и грибами, повреждении брухусом и гороховой плодожоркой, а также выходе колотого зерна при лущении.
Принципиальная возможность этого направления селекции доказана. Новый мелкосемянный сорт Мультик (масса 1000 семян 140 г) в среднем за три года конкурсного испытания показал практически одинаковую урожайность с крупносемянным (масса 1000 семян 250 г) сортом Орловчанин, но содержал белка в среднем на 1,5% больше.
За период селекции от примитивных форм до лучших современных сортов накопление белка в семенах уменьшилось на 2–7% (в абсолютном выражении). Основной причиной этого является увеличение доли зерна в биомассе при неизменившемся фонде азота в растении, в результате чего на один грамм семян стало приходиться на 22...38% меньше азота.
Главный путь увеличения белковости семян гороха методами селекции – это скрещивания по схеме высокобелковый х высокобелковый. И все же объединить в генотипе высокую белковость и урожай будет непросто ввиду существования между ними отрицательной корреляции. Исходя из этого, обоснованным является создание высокоурожайных сортов при среднем содержании белка в семенах, учитывая и другие их потребительские признаки: цвет семенной кожуры, пригодность к лущению и кулинарной обработке. Вместе с тем, по данным французских исследователей (Natalie-Munier et al., 2009), подкормка азотными удобрениями в начале налива семян может исключить отрицательную корреляцию между урожайностью и содержанием белка в семенах.
Проведенный выше ретроспективный анализ формирования урожая на примере кукурузы, пшеницы, сои и гороха отчетливо показывает, что основную роль в реализации потенциала урожайности играют преимущественно экстенсивные, а не интенсивные, в частности ИФ, параметры. Тем не менее роль последней становится определяющей в фазе генеративного развития, когда устойчивость процессов фотоассимиляции двуокиси углерода и налива семян может решить судьбу урожая. Поэтому анализ донорно-акцепторных отношений а также создание сортов с повышенной аттрагирующей способностью плодоэлементов в генеративную фазу развития и высокой потенциальной урожайностью для адаптивной системы земледелия, устойчивых к широкому спектру биотических и абиотических стрессоров, остается главной задачей селекционеров.
1.2. Сигналинг растений в условиях стресса
Воздействие абиотических (засуха, засоление, холод, мороз, тяжелые металлы и др.) и биотических (патогены) стрессоров на растения приводит к целому ряду неспецифических ответных реакций, что и позволило называть его адаптационным синдромом, или стрессом. Указанные выше реакции являются результатом «включения» стрессорами неспецифических сигнальных систем в клетке.
Молекулярные механизмы регуляции клеточного метаболизма внешними (первичными) сигналами, несущими в клетку информацию, что принципиально отличает их от других поступающих в клетку химических соединений, служащих для нее источником материи и энергии, являются предметом исследования «клеточной сигнализации». Однако последняя подразумевает не только передачу сигналов как таковую, но и весь комплекс событий, с ней сопряженных, в том числе усиление (или амплификацию), ослабление (или аттенюацию) и подавление (или выключение) сигналов (Филиппов, 1997).
Звенья сигнальных систем. Сигналы, поступающие из окружающей среды, улавливаются клеткой с помощью специальных «антенн» – рецепторных белковых молекул, пронизывающих клеточную мембрану и выступающих над ее поверхностями с наружной и внутренней стороны. Нековалентное взаимодействие внешнего участка рецептора с той или иной сигнальной молекулой, поступающей из среды, окружающей клетку, приводит к изменению конформации рецепторного белка, которое передается на внутренний, цитоплазматический участок. В большинстве сигнальных систем с ним контактируют посреднические G-белки – еще одно унифицированное (по своим структуре и функциям) звено сигнальных систем. G-белки выполняют функции преобразователя сигналов, передавая сигнальный конформационный импульс на стартовый фермент, специфичный для той или иной сигнальной системы. Еще одним звеном сигнальных систем являются протеинкиназы (ферменты, переносящие концевой остаток ортофосфорной кислоты с АТФ на те или иные белки), активируемые продуктами стартовых сигнальных реакций или их производными. Фосфорилированные с помощью протеинкиназ белки являются следующими звеньями сигнальных цепей.
Еще одно унифицированное звено сигнальных систем клеток – это белковые факторы регуляции транскрипции, которые представляют собой один из субстратов протеинкиназных реакций. Структура этих белков также в значительной степени унифицирована, а модификации структуры определяют принадлежность факторов регуляции транскрипции к той или иной сигнальной системе. Фосфорилирование факторов регуляции транскрипции обусловливает изменение конформации этих белков, их активацию и последующее взаимодействие с промоторным участком определенного гена, что приводит к изменению интенсивности его экспрессии (индукции или репрессии), а в крайних случаях – к «включению» некоторых молчавших генов или «выключению» активных. Репрограммирование экспрессии совокупности генов генома вызывает изменение соотношения белков в клетке, что и является основой ее функционального ответа. В отдельных случаях химический сигнал из внешней среды может взаимодействовать с рецептором, расположенным внутри клетки – в цитозоле или даже ядре. Однако поскольку информационный путь имеет меньшее число интермедиатов, у него и меньше возможностей для регуляции со стороны клетки.
Внешняя среда, рецепторы, мессенджеры. Разнообразные молекулы, инициирующие трансмембранную передачу сигналов, активируют рецепторы, действуя на них обычно при очень низких концентрациях, порядка 10–8 М и ниже. Активированный рецептор тем или иным способом передает сигнал к внутриклеточным мишеням. Если мишень, или эффекторный белок, представлена ферментом, то сигнал модулирует (увеличивает или уменьшает) его каталитическую активность; если эффекторным белком служит ионный канал, то модулируется проводимость этого канала. В обоих случаях результатом будет изменение активности какой-то метаболической стадии (стадий), либо цитоплазматической концентрации того или иного иона и, как следствие, возникновение клеточного ответа.
Молекулярные машины, обеспечивающие передачу сигнала от рецепторов к внутриклеточным мишеням, состоят, как правило, из нескольких белковых компонентов, совокупность которых обычно именуют каскадом передачи сигнала или просто каскадом. Помимо белковых посредников в передачу сигнала внутри клетки во многих случаях вовлекаются и относительно небольшие молекулы, служащие вторичными сигналами, – это мессенджеры (от англ. messenger – посыльный). Дело в том, что в цитоплазме в передачу сигнала вовлечены как разнообразные белки, так и малые молекулы (собственно вторичные сигналы), причем функционально все они являются посредниками между рецептором, на который подействовал внешний стимул, и клеточным ответом. Однако между ними есть и принципиальное различие: белки образуют своеобразную молекулярную машину, которая, с одной стороны, чувствует внешний сигнал, а с другой – обладает ферментативной или иной активностью, модулируемой этим сигналом, в то время как малые молекулы действительно служат посыльными (мессенджерами) между различными белками, полиферментными комплексами или даже клеточными структурами.
Наиболее универсальными вторичными мессенджерами являются ионы кальция (Са2+), циклический АМФ (цАМФ), циклический ГМФ (цГМФ), диацилглицерол (ДАГ), инозитолтрифосфат (ИТФ). Вторичные мессенджеры значительно усиливают первичный сигнал. В результате одна-единственная молекула гормона может привести к активации фермента, продуцирующего многие сотни молекул вторичных мессенджеров.
В норме концентрация вторичных мессенджеров в клетке ничтожно мала. В ответ на связывание гормона или другое раздражение активированный рецептор передает сигнал на ферментные системы образования одного или нескольких вторичных мессенджеров. Концентрация мессенджеров в цитоплазме резко возрастает, после чего возвращается к низкому исходному уровню. Кратковременный «всплеск» уровня вторичного мессенджера в клетке оказывается достаточным для активации или инактивации ферментов и регуляторных белков, инициирующих ответ организма на сигнал.
Ионы кальция. В растительных клетках Са2+ содержится в клеточных стенках в форме пектатов, карбонатов, сульфатов, но большинство – в вакуоли в форме оксалата благодаря работе Ca2+/H+ антипорта тонопласта. Его концентрация в цитозоле в нормальных условиях крайне низка и составляет около 1×10–7 М, но заметно выше – до 1 ‧ 10–3 М в митохондриях и ЭПР. Такой огромный градиент поддерживается благодаря работе, расположенной на плазмалемме Са2+-АТФазе, которая выбрасывает Са2+ из цитозоля в свободное пространство клеточной стенки. Вторая Са2+-АТФаза, также снижающая концентрацию Ca в цитозоле, находится на мембранах ЭПР и транспортирует кальций в цистерны из цитозоля.
В ответ на действие различных факторов открываются кальциевые каналы, и концентрация Са2+ в цитозоле возрастает в течение секунд или минут. Однако подобная динамика носит обратимый характер.
Са2+ может связываться с регуляторным белком кальмодулином, имеющим 4 участка с высоким сродством к Са2+ и содержащимся в митохондриях, хлоропластах, клеточных стенках. Связывание кальмодулина с Са2+ изменяет конформацию его молекулы, что придает этому комплексу способность активировать некоторые ферменты: Са2+-АТФазы, НАД-киназы, НАД-оксидоредуктазы, протеинкиназы, липазы.
Протеинкиназы – ферменты, фосфорилирующие белки (структурные, транспортные и регуляторные) по строго определенным группам серина, треонина и тирозина, что приводит к изменению структуры белковой молекулы и ее функциональной активности. Активированная протеинкиназа переносит фосфатную группу с АТФ на белки, которые активируют другие ферменты. Биологический смысл этой цепи реакций состоит в том, что усиливается первичный сигнал, вследствие чего включается синтез стрессорных белков, защитных соединений (сахарозы, пролина, олигосахаров), меняется липидный и белковый состав мембран.
Протеинкиназы активируются кальцием, фосфолипидами (фосфатидилсерином) и диацилглицеролом. В растениях функционирует Са2+ – зависимая протеинкиназа (CDPK), участвующая в трансдукции сигнала. Причем одна из субъединиц этого фермента представляет собой кальмодулин, который способен связывать ионы кальция. Большинство ответных реакций, вызываемых Са2+, обусловлены активацией этой протеинкиназы (CDPK). Активированная протеинкиназа переносит фосфатную группу с АТФ на белки.
В клетке существует серия протеинкиназ, которые последовательно фосфорилируют друг друга, в результате происходит «каскад» реакций. Этот каскад был назван МАРК-каскадом от митоген-активируемой протеинкиназы (англ. – Mitogen Activated Protein Kinese, или МАРК). Первая киназа в этой последовательности ферментов называется МАРККК (МАР-киназа-киназа киназы). Именно она «запускает» весь МАРК-каскад. МАРККК передает фосфорный остаток другой киназе – МАРКК (МАР-киназа киназы), которая, в свою очередь, передает сигнал МАР-киназе (МАРК). Она направляется к ядру, где активирует другие протеинкиназы и различные транскрипционные факторы, которые связываются с цис-действующими регуляторными элементами и вызывают дифференциальную экспрессию генома. МАРК-каскад является важным способом передачи (трансдукции) внутриклеточного сигнала у растений (Кузнецов, Дмитриева, 2005).
Циклические мононуклеотиды. Помимо ионов кальция наиболее универсальными вторичными мессенджерами являются циклические мононуклеотиды. У животных циклический аденозинмонофосфат (цАМФ) и циклический гуанозинмонофосфат (цГМФ) играют важную роль в регуляции клеточных процессов. По своему строению цАМФ похож на АТФ, образуется из АТФ в результате отделения двух остатков фосфорной кислоты и последующего замыкания оставшейся фосфатной группировки в кольцо. Отсюда и произошло название мононуклеотида. Эта реакция катализируется аденилатциклазой, которая расположена на внутренней поверхности мембран и работает в присутствии фосфолипидов и ионов магния. В растениях концентрация цАМФ крайне низка. Циклический аденозинмонофосфат активирует многие ферменты. В последние годы накапливается все больше данных в пользу того, что цАМФ в растительных клетках выполняет столь же важную сигнальную роль, как в клетках животных и прокариот.
Циклический АМФ действует как аллостерический эффектор по отношению к протеинкиназам, которые фосфорилируют ядерные, цитоплазматические и мембранно-связанные белки.
В последнее время появляются доказательства того, что циклический гуанозинмонофосфат (цГМФ) тоже функционирует в качестве вторичного мессенджера в растениях, подобно животным. В животных клетках цГМФ регулирует ионные каналы и протеинкиназы.
Основные типы мембранных рецепторов. Существуют три основных типа рецепторов, интегрированные во внешнюю клеточную мембрану: 1) рецепторы, сопряженные с G-белками, 2) рецепторы – ионные каналы; 3) рецепторы, ассоциированные с ферментативной активностью.
Рецепторы, сопряженные с G-белками (их сокращенное обозначение GPCR, от G-protein coupled receptors), передают сигнал от первичных мессенджеров к внутриклеточным мишеням с помощью каскада GPCR ⇒ G-белок ⇒ эффекторный белок. Первичными сигналами для этих рецепторов служат самые разнообразные молекулы, среди которых в животных клетках низкомолекулярные гормоны и нейропередатчики, или нейротрансмиттеры (например, адреналин, ацетилхолин, норадреналин и др.). Некоторые из них, кстати, обнаружены и в растениях.
Один и тот же первичный сигнал может инициировать передачу сигнала через несколько (иногда более 10) разных GPCR, так что если число внешних сигналов для GPCR составляет несколько десятков, то самих таких рецепторов известно более 200. При всем их разнообразии GPCR представляют собой мономерные интегральные мембранные белки, полипептидная цепь которых семь раз пересекает клеточную мембрану. Во всех случаях участок рецептора, ответственный за взаимодействие с первичным сигналом, локализован с внешней стороны мембраны, а участок, контактирующий с G-белком, – на ее цитоплазматической стороне.
Следующий за рецептором «вниз по течению» компонент каскада передачи сигнала с участием GPCR представлен G-белком. Найдено около 20 различных G-белков.
Важнейшая характеристика G-белков – присутствие на их α-субъединице центра связывания гуаниловых нуклеотидов: ГДФ и GTR. Если с G-белком связан ГТФ, то это соответствует его активированному состоянию (G-ГТФ), или, иначе, G-белок находится в положении «включено». Если в нуклеотидсвязывающем центре присутствует ГДФ, то эта форма (G-ГДФ) соответствует состоянию «выключено». Центральное событие при передаче сигнала от рецептора, на который подействовал первичный сигнал, к G-белку состоит в том, что активированный рецептор катализирует обмен ГДФ, связанного с G-белком, на присутствующий в среде ГТФ. Это событие, обозначаемое как ГДФ/ГТФ-обмен на G-белке, сопровождается диссоциацией тримерной молекулы G-белка на две функциональные субъединицы: α-субъединицу, содержащую ГТФ, и δγ-комплекс. Далее одна из этих функциональных субъединиц, какая именно – зависит от типа сигнальной системы, взаимодействует с эффекторным белком, представленным ферментом или катионным каналом. Как следствие, их каталитическая активность или ионная проводимость соответственно меняется, что, в свою очередь, приводит к изменению цитоплазматической концентрации вторичного мессенджера (или катиона) и, в конечном счете, инициирует тот или иной клеточный ответ.
Весьма важно, что при передаче сигнала в каскаде рецептор ⇒ G-белок ⇒ эффекторный белок исходный внешний сигнал может многократно усиливаться, или амплифицироваться. Это происходит благодаря тому, что одна молекула рецептора за время пребывания в активированном состоянии (R*) успевает перевести в активированную форму (G*) несколько молекул G-белка. В конечном счете, коэффициент амплификации сигнала при его прохождении через каскад может достигать весьма высоких значений: в зрительных клетках это величина порядка 105–106.
Ясно, что прекращение действия внешнего стимула должно сопровождаться «выключением» всех компонентов сигнальной системы. На уровне рецепторов это достигается, во-первых, в результате диссоциации первичного сигнала из комплекса с GPCR, во-вторых, путем фосфорилирования рецепторов под действием специальных протеинкиназ и последующего связывания с модифицированным рецептором специального белка.
Поскольку состояние активации эффекторного белка (включен–выключен) прямо зависит от состояния G-белка, то этот переход означает также выключение эффекторного белка, а следовательно, прекращение синтеза (гидролиза) вторичного мессенджера или закрывание ионного канала. И, наконец, чтобы переход клетки к исходному (до действия внешнего стимула) состоянию завершился, специальные механизмы восстанавливают исходный уровень вторичного мессенджера или катиона в ее цитоплазме.
Рецепторы – ионные каналы – это интегральные мембранные белки, состоящие из нескольких субъединиц, полипептидная цепь которых несколько раз пересекает наружную клеточную мембрану. Они действуют одновременно и как ионные каналы, и как рецепторы, которые способны специфически связывать со своей внешней стороны первичные сигналы, изменяющие их ионную (катионную или анионную – в зависимости от типа рецептора) проводимость.
Рецепторы, ассоциированные с ферментативной активностью, по своей четвертичной (субъединичной) структуре весьма разнообразны. За некоторыми исключениями они представляют собой либо мономеры, которые при связывании с ними первичного мессенджера димеризуются, либо олигомеры, образованные несколькими субъединицами различных типов. Практически у всех этих рецепторов полипептидная цепь их мономерных субъединиц единственный раз пересекает клеточную мембрану. Общим у них является также то, что участок для связывания первичного сигнала локализован на рецепторе со стороны, обращенной во внеклеточное пространство.
В настоящее время интенсивно исследуются МАР-киназная, аденилатциклазная, фосфатидатная, кальциевая, липоксигеназная, НАДФН-оксидазная, NO-синтазная и протонная сигнальные системы и их роль в онтогенетическом развитии растений и в формировании ответа на изменяющиеся условия существования, особенно на действие различных абиотических и биотических стрессоров.
Контрольные вопросы и задания
- Что понимается под клеточной сигнализацией?
- Какие вещества функционируют в качестве рецепторов в клеточной сигнализации?
- Какова роль G-белков в каскаде передач сигнала?
- Какую роль выполняют вторичные мессенджеры?
- За счет чего изменяется концентрация вторичных мессенджеров в ответ на раздражение?
- Какие низкомолекулярные вещества выступают в роли вторичных мессенджеров?
- Какова роль Ca-каналов в передаче сигнала?
- Какой блок обеспечивает мессенджерную роль кальция?
- Какова роль протеинкиназ в функционировании сигнальных систем?
- Как функционирует МАРК-каскад в клетке?
- Опишите возможные физиологические подходы по повышению потенциальной продуктивности посевов сельскохозяйственных культур.
- Почему урожайность зернобобовых культур кратно уступает урожайности кукурузы?
- В чем причина большой разницы между потенциальной и реальной урожайностью культур?
- Назовите возможные пути повышения КПД ФАР посевов.
- Дайте ретроспективную оценку вклада селекции и агротехнологий в повышение урожайности полевых культур.
- Возможно ли в ближайшей перспективе повышение эффективности трансформации ФАР в биомассу с использованием трансгенных технологий?
Глава 2. ОКИСЛИТЕЛЬНЫЙ СТРЕСС
2.1. Активные формы кислорода
2.1.1. Характеристика основных форм кислорода
Термин «активные формы кислорода» обозначает совокупность короткоживущих взаимопревращающихся реакционноспособных форм кислорода, возникающих в результате его электронного возбуждения или окислительно-восстановительных превращений. Основное количество кислорода (95–98%) у растений расходуется в качестве универсального конечного акцептора электронов при дыхании, а также при окислении различных субстратов. В активные формы кислорода (АФК) переходит относительно небольшая, но высоко агрессивная часть кислорода (2–5%), способная повреждать практически все компоненты клетки.
Они образуются в организме в реакциях одно-, двух- и трех-электронного восстановления кислорода в результате само- и ферментативного окисления соединений, а также в фотоиндуцированных реакциях. Среди АФК, кроме свободно-радикальных частиц, обозначаемых символом « • » – супероксидного анион-радикала (O2–), гидроксильного радикала (ОН·), перекисных радикалов (RO2·) и др., – различают и нейтральные молекулы, такие как пероксид водорода (Н2О2), синглетный кислород (1O2), озон (О3) и др.
Полное восстановление молекулы кислорода до воды в дыхательной цепи требует четырех электронов, формирование же АФК происходит при неполном одноэлектронном восстановлении, в результате которого образуются свободные радикалы:
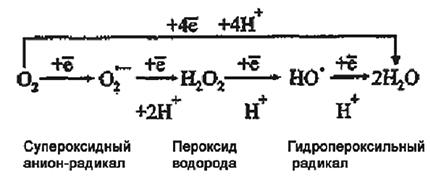
Супероксидный анион-радикал O2 является первичным продуктом восстановления, как и его протонированная форма, гидропероксидный радикал (НO2·) при слабокислых значениях рН. Супероксидный анион-радикал локализуется во внутренней мембране митохондрий вблизи от митохондриальной ДНК, которая становится для него наиболее уязвимой мишенью. O2– не обладает сильными окислительными свойствами, но представляет большую опасность, поскольку является источником образования более реакционноспособных АФК. В отличие от молекулярного кислорода, для передвижения которого мембраны не представляют преграды, супероксидный анион, обладая зарядом, окружен молекулами воды и не может преодолеть гидрофобный мембранный барьер. Кроме того, он имеет большое время жизни и становится источником других АФК.
В результате присоединения одного электрона и двух протонов к супероксидному радикалу или двухэлектронного переноса на молекулу O2 образуется пероксид водорода (Н2O2 ), который относится к окислителям средней силы и служит источником очень опасного гидроксильного радикала. Н2O2 возникает в ферментативных реакциях с оксидазами, переносящими 2е– на О2, и реакции дисмутации НO2· и O2· в основном с участием, а также и без участия супероксиддисмутазы (СОД):
2 НO2· → Н2O2 + O2
НO2· + O2· + Н+ → Н2O2 + O2
2 O2· + 2Н+ → Н2O2 + O2
Перекись водорода выполняет в растительной клетке следующее: активирует (или репрессирует) работу генов множества защитных ферментных белков, сигнальных белков – МАР-киназ и протеинфосфатаз, белков, регулирующих окислительный статус клетки (НАДФН оксидазы и ОхуR белка), а также активирует Са-каналы и вызывает закрывание устьиц, участвует в процессах лигнификации и в индукции апоптоза совместно с NO, является сигнальной молекулой-переносчиком стрессорного сигнала.
В отсутствие металлов переменной валентности пероксид достаточно стабилен. В то же время благодаря относительно продолжительному времени жизни и растворимости в липидном бислое пероксид может легко диффундировать через мембраны. Роль пероксида определяется не столько собственной химической реактивностью, хотя он и не инициирует перекисное окисление липидов (ПОЛ), сколько участием в образовании более токсичных АФК.
При дальнейшем одноэлектронном восстановлении возможно появление гидроксильного радикала НО·, очень сильного окислителя. Он не способен к внутриклеточной миграции, так как моментально вступает в реакцию с биомолекулами. Основным источником НО· служит реакция Фентона с участием металлов, главным образом Fe2+: Fe2+ + H2O2 → Fe3+ + ОН– + НО·. Металлы участвуют в реакции генерации НО· не только в составе неорганических солей, но и в составе белков.
Гидроксильный радикал вызывает окисление пуриновых и пиримидиновых оснований нуклеиновых кислот, их модификацию, разрывы цепей, повреждение хромосом. По-видимому, АФК обладают большим мутагенным действием, чем другие классы мутагенов.
Гидроперекисный анион (НO2–) и пероксидный анион (O22–) – продукты двухэлектронного переноса на молекулу O2. Радикалы O22– и НO2– быстро протонируются в кислой среде, но могут вступать в реакции с органическими радикалами. НО2– – более сильный окислитель, чем O2–, и может вызывать перекисное окисление липидов, что, вероятно, обусловлено его большей гидрофобностью.
Оксильный анион-радикал (O–) и гидроксильный радикал (ОН·) – продукты трехэлектронного восстановления. При физиологических значениях рН O– существует в виде ОН·, который является наиболее реакционноспособной формой АФК (время жизни 10–9 с). Он разрушает практически любую встретившуюся+ ему молекулу и инактивирует ферменты. В нуклеиновых кислотах ОН· разрушает углеводные мостики между нуклеотидами, разрывая цепи ДНК и РНК. Внедряясь в липидный слой, радикал гидроксила запускает реакции перекисного окисления, что приводит к повреждению мембран.
Синглетный кислород (1O2) генерируется при изменении спина одного из электронов, находящихся на π-орбиталях O2. Это одна из наиболее реакционно-способных форм активированного кислорода, окисляющего большинство органических молекул с образованием гидроперекисей (RH + O2 → ROOH). Cинглетный кислород в основном синтезируется в фотоиндуцированных реакциях при участии хлорофиллов, других порфиринов, флавинов. Образование 1O2 возможно также в реакциях дисмутации O2·, НO2·, ОН· и при работе ферментов (СОД, каталазы, пероксидазы).
АФК могут образоваться также при взаимодействии озона с O2, в результате чего возникает супероксид-анион: O3 + O2→ 2O2 + O2·, а затем – пероксид водорода, синглетный кислород и гидроксильный радикал.
Взаимодействуя с органическими веществами, АФК, главным образом НО·, образует гидропероксиды (RООН) ДНК, белков, липидов. Образование ROОН называют перекисным окислением (пероксидацией). В липидах (L), в основном в полиненасыщенных жирных кислотах, АФК вызывают цепные реакции с накоплением липидных радикалов (L·), пероксилов (LOO·), гидропероксидов (LOOH) и алкосилов (LO·):
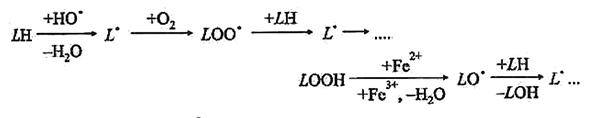
Реакция LOOH с Fe2+ ведет к разветвлению цепи. Далее образуются диеновые (триеновые) коньюгаты жирных кислот, а конечными продуктами являются минорные метаболиты – малоновый диальдегид, этан, пентан и др. Этот процесс, называемый перекисным окислением липидов (ПОЛ), может происходить спонтанно, неферментативно, но большое значение имеют и ферментативные реакции, например с участием липоксигеназ.
Реакции ПОЛ мембран хлоропластов, причем преимущественно ненасыщенных жирных кислот (НЖК), включают также взаимодействие с белковыми молекулами мембран. Основным типом изменения белков является окисление их сульфгидрильных групп. В результате образуются сульфгидрильные радикалы, которые затем взаимодействуют с образованием дисульфидов либо окисляются кислородом с образованием производных сульфоновой кислоты.
В результате происходит инактивация ферментов, а также отмечается появление дефектов в липидном слое мембран и соответственно увеличение ионной проницаемости липидного бислоя и утечка электролитов. Кроме того, уменьшается стабильность мембран, особенно сопрягающих мембран (хлоропластов и митохондрий) и плазмалеммы. Таким образом ПОЛ нарушает процессы фото- и окислительного фосфорилирования, синтез АТФ в митохондриях и хлоропластах, и клетка оказывается в условиях энергетического голода. Одновременно в цитоплазму выходят ионы Са2+ (см. гл. 1). Активация ПОЛ на плазмалемме также вызывает нарушение ее функций, приводит к изменению всей клеточной жизнедеятельности или даже к гибели клеток.
Продукты взаимодействия производных кислорода – окиси азота (NO) и особенно его радикала (NO·) с O2 вызывают окисление аминов, тиолов и других молекул. Взаимодействие NO· с O2– приводит к генерации крайне токсичного и мощного оксиданта – пероксинитрита (ONOО–), а также ОН·.
2.1.2. Биологическое значение АФК
В нормальных условиях жизни растений образование АФК и ПОЛ – обычные метаболические процессы. Патологические последствия возникают при чрезмерном накоплении АФК, пероксидов и их производных в результате стрессовых воздействий. Концентрация АФК в условиях стресса может возрасти в 3–10 раз (Polle, 2001).
Так, генерация АФК может быть необходима для формирования защитных реакций растений, например при инфекционных заболеваниях. АФК, локализованные на поверхности клетки – в плазматической мембране и клеточной стенке, участвуют в метаболизме фенольных соединений, в частности в синтезе лигнина, обеспечивающего механическую прочность клеточной стенки. Образующиеся в матриксе клеточной стенки кислородные радикалы способны атаковать и повреждать патогенную микрофлору. Таким образом создается внеклеточный защитный барьер, препятствующий проникновению патогенных микроорганизмов внутрь растительной клетки.
Образование АФК – наиболее ранний ответ растения-хозяина на инфекцию, который позволяет «узнать» патогена. Исследования взаимоотношений патогена с растением-хозяином выявили также сигнальную и регуляторную функции АФК, включающие другие, более поздние защитные реакции в клетках растений. Именно этим функциям АФК уделяется основное внимание в последние годы. В низких субтоксических концентрациях АФК и органические гидропероксиды индуцируют экспрессию генов и деление клеток. АФК могут служить вторичными посредниками в передаче сигнала от ЭТЦ митохондрий геному ядра. Так, повышение уровня АФК в митохондриях в присутствии ингибиторов цитохромного пути и салициловой кислоты, ингибирующей каталазу, индуцировало экспрессию гена АОХ I, кодирующего синтез альтернативной оксидазы, а также синтез белков, определяющих устойчивость к патогенам.
АФК вызывает повышение концентрации Са2+ в цитозоле и стимуляцию фосфорилирования белков в результате активации протеинкиназ (особенно протеинкиназы С), протеинтирозинкиназ и ингибирования протеинфосфатаз.
Условия образования и роль АФК в растительных клетках. АФК образуются при нормальном метаболизме клетки. Супероксидный радикал и синглетный кислород обычно продуцируются в освещенных хлоропластах путем сопутствующего переноса электронов от возбужденной молекулы хлорофилла или компонентов ФСI при условии высоких отношений НАДФН/НАДФ к молекулярному кислороду. Каротиноиды эффективно гасят возбужденное триплетное состояние хлорофилла и/или синглетного кислорода.
В генерировании АФК значительную роль играет озон, образующийся в результате фотодиссоциации молекулярного кислорода под влиянием солнечной энергии. При взаимодействии озона с O2 возникает супероксид-анион: O3 + O2 → 2O2 + O2–, а затем – пероксид водорода, синглетный кислород и гидроксильный радикал. Озон связывается с плазмалеммой клеток листьев, способствует нарушению регуляции устьичного аппарата, повреждению тилакоидных мембран, деградации (фотоингибированию) РБФ-карбоксилазы и торможению фотосинтеза в целом (см. гл. 13).
АФК участвуют в регуляции липидного состава и проницаемости мембран, устьичных движений, способствуют синтезу многих веществ, в частности, фитогормонов и физиологически активных соединений – этилена, салициловой кислоты, жасмоната и других. Выявлено их значение для синтеза клеточной стенки и кросс-связывания, полимеризации лигнина и суберина. Механизм фотовосстановления протохлорофиллида также включает стадию образования свободных радикалов. Локальный окислительный взрыв (проявляющийся в накоплении Н2О2) участвует в контроле проникновения образующих арбускулярную микоризу грибов в клетки корня.
АФК участвуют в поддержании окислительно-восстановительной среды клетки и тиолового статуса белков, тем самым регулируя многочисленные процессы (Schafer, Buetter, 2001). Предполагается, что они контролируют митоз, дифференцировку пластид, аппарата Гольджи (Бакеева и др., 2001) и необходимы для нормального роста, развития и морфогенеза растений (Шорнинг и др., 2000; Нариманов, 2001).
ПОЛ на определенном уровне совершенно необходимо для обновления фосфолипидов мембран, регуляции транспорта через мембрану, синтеза различных веществ (спирты, кетоны, альдегиды, диальдегиды, эпоксиды, полимеры, фураны и др.). Дальнейшее окисление первичных продуктов перекисного окисления – гидропероксидов – может привести к образованию физиологически активных соединений, в частности, этилена, цитокининов, жасмоновой кислоты и др.
Вышесказанное свидетельствует о заметной роли АФК в нормальном метаболизме клеток. Однако действие разных стрессоров* (УФ-свет и другие формы радиации, гербициды, патогены, механические повреждения, аноксия, озон, температурный, солевой и другие стрессы) может приводить к избытку АФК, превышающему возможности системы гашения с помощью антиоксидантов, что обуславливает необходимость добавочной защиты. Например, закрывание устьиц в условиях засухи ограничивает доступность СО2 для фотосинтетической ассимиляции углерода. В этих условиях, как и на интенсивном солнечном свету, избыточное образование супероксида в хлоропластах может приводить к фотоингибированию и фотоокислению.
Все вышеприведенные воздействия на растения имеют обратную связь с хлоропластом. Независимо от метаболического участка в растении, на который влияет какой-либо стрессор, всегда происходят фотосинтетическая активация кислорода и/или фотодинамические процессы, приводящие к образованию АФК.
Одной из главных мишеней действия АФК в стрессовых условиях является ДНК. При этом повреждаются основания и дезоксирибоза, а также появляются ковалентные сшивки и разрывы. Предполагается, что в разрушении молекулы участвует преимущественно гидроксильный радикал, в то время как супероксид и пероксиды практически не реагируют с ДНК. Окисление нуклеиновых кислот возможно не только путем прямого действия АФК, но и через активацию ими Са2+-зависимых эндонуклеаз.
Компартментация АФК. Хлоропласт. Супероксидный анион-радикал возникает непосредственно в ходе работы фотосинтетической ЭТЦ (реакция Мелера). Основным участком фотовосстановления O2 в хлоропластах является ФСI, а главным восстановителем считается ферредоксин (Fd) (или ферредоксин-НАДФН-редуктаза). Установлено, что в ФСI образуется в основном O2–, в то время как в ФСII – O2– и Н2O2 (Tjus et al., 2001).
В хлоропластах происходит также активное образование синглетного кислорода 1O2, возникающего за счет взаимодействия O2 с возбужденным хлорофиллом (Красновский, 1994). 1O2 легче вступает в реакции с другими молекулами, чем кислород в обычном состоянии. По некоторым данным, 1O2 является основным инициатором ПОЛ в тилакоидных мембранах (Иванов, 1998). Генерация АФК в хлоропластах происходит также в темновых реакциях при участии РБФК.
Митохондрии. Они являются минорными участками активации кислорода и способны продуцировать O2– и пероксид водорода при окислении различных субстратов, причем большая часть Н2O2 образуется в реакции дисмутации O2–. Возникновение супероксида происходит в цепи дыхательных переносчиков, в основном за счет убихинона, флавинов и Fe-содержащих белков. В ходе окислительного фосфорилирования при нормальных условиях около 5% кислорода преобразуется в АФК; их генерация существенно увеличивается при снижении концентрации АДФ, поскольку из-за уменьшения потребления кислорода возрастает его концентрация и повышается степень восстановленности переносчиков в ЭТЦ.
Плазмалемма. Плазматические мембраны содержат НАД(Ф)Н-оксидазы, которые относительно специфичны к своим акцепторам электронов и переносят электроны с цитоплазматического НАД(Ф)Н на кислород апопласта. Супероксид и его производные наиболее интенсивно продуцируются в присутствии автоокислительных акцепторов, таких как хиноны; в данном случае АФК могут функционировать в качестве клеточных мессенджеров.
Значительную роль в образовании АФК и активации процесса ПОЛ на плазмалемме играют липоксигеназы, которые катализируют образование гидроперекисей из ненасыщенных жирных кислот путем присоединения к ним молекулы O2. Указанные гидроперекиси могут легко распадаться с образованием свободных радикалов. В некоторых случаях возникновение RO2·, O2– и других АФК обнаружено в самой липоксигеназной реакции. Все эти радикалы являются источниками окислительного повреждения мембраны (Veronesi et al., 1996).
Микросомы. Микросомы образованы из ЭПР и содержат две системы транспорта электронов. Одна из них – НАДФ-зависимая система – способна восстанавливать O2 по одно-, двух- и трехэлектронному механизму (Дмитриев, 1998). Возникновение O2– обусловлено цитохромом P450, цитохромом bs и НАД(Ф)Н-цитохром b5(с)-редуктазой.
Микротельца (пероксисомы и глиоксисомы). Микротельца, в отличие от митохондрий или хлоропластов, являются компартментами с одиночной мембраной, содержащей ксантиноксидазу, которая образует О2– и Н2О2, а также гликолатоксидазу и β-оксидазу жирных кислот, продуцирующие Н2О2. Регуляторную роль может играть ингибирование ксантиноксидазы некоторыми флавоноидами.
В глиоксисомах функционирует оксидазная форма ксантиноксидазы. В пероксисомах образование АФК происходит в ходе β-окисления жирных кислот, энзиматических реакций флавиновых оксидаз и гликолатоксидазной реакции фотодыхания. В матриксе органелл находится ксантиноксидаза, в определенных условиях генерирующая большие количества О2–.
Клеточная стенка. В генерации АФК апопласта (O2– и Н2O2) принимает участие пероксидаза клеточной стенки, продуцирующая O2– при расходовании НАДН в Мn2+-зависимой реакции. НАДФН-оксидаза отвечает за поставку H2O2 для пероксидаз в процессах лигнификации клеточной стенки. Кроме того, в клеточной стенке найдены аминооксидазы, продуцирующие Н2O2.
Определенный вклад в генерацию АФК в клетке вносит ядерная мембрана, в которой О2– образуется за счет окисления НАДФН. Образование АФК возможно в цитозоле при автоокислении и ферментативных реакциях с участием каталазы, пероксидазы и других ферментов.
2.1.3. Механизмы детоксикации АФК
На сегодняшний день известны следующие методы определения уровня и локализации окислительного стресса в растениях:
• определение активности антиоксидантных ферментов;
• определение концентрации АФК: супероксид-ионов, перекиси водорода и др.;
• измерение концентрации маркеров окислительного стресса: малонового альдегида (ПОЛ), окисленного гуанозина и др.;
• гистохимические методы определения локализации АФК;
• прямое измерение концентрации АФК методом парамагнитного резонанса;
• масс-спектрометрические методы измерения количества АФК в клетке.
Классификация антиоксидантов. Под антиоксидантами (АО) понимают широкий класс веществ, в которую входят соединения, снижающие активность радикальных окислительных процессов по следующим механизмам:
• ингибирование радикальных форм активных кислородных метаболитов, способных инициировать образование органических радикалов;
• изменение структурной организации субстрата, замедляющее окисление;
• прерывание окислительной цепи посредством взаимодействия с органическими радикалами;
• снижение концентрации кислорода;
• связывание или окисление ионов металлов переменной валентности, инициирующих разложение перекисей и образование радикалов;
• перевод перекисей в стабильные продукты окисления: спирты, альдегиды, кетоны и др.
Выделяют также антиоксиданты вторичной защиты, которые обеспечивают восстановление от повреждений из-за действий АФК. Сюда относятся процессы репарации ДНК, гидролитические ферменты, осуществляющие «выщепление» окисленных липидов из мембраны, протеолиз поврежденных белков, комплекс реакций, связанных с метаболизацией продуктов ПОЛ. Детоксикация в широком смысле также включает замещение поврежденных молекул, таких как ДНК, белки и мембранные липиды, комплексной системой интегрированных репарирующих ферментов и процессов замещения. Однако непрерывная работа этих процессов репарации приведет к их инактивации, так как они функционируют также и в качестве мишеней АФК.
Наиболее удобно характеризовать группы антиоксидантов в зависимости от их молекулярных масс, причем первую группу составляют низкомолекулярные соединения, а вторую – высокомолекулярные ферменты, белки и пептиды, способные связывать ионы металлов переменной валентности.
Низкомолекулярные антиоксиданты. К ним относятся разнообразные соединения – глутатион, аскорбат, токоферолы, каротиноиды, полиамины, некоторые аминокислоты и др. Отличительной особенностью большинства эндогенных низкомолекулярных антиоксидантов является нелинейная зависимость между их концентрацией и степенью ингибирования свободнорадикальных процессов. Антиоксиданты (АО) можно подразделить на две группы: водорастворимые протектанты и антиоксиданты липидной фазы.
Из водорастворимых антиоксидантов наиболее эффективными являются глутатион и аскорбиновая кислота, обнаруженные практически во всех участках клетки, но в особенно больших концентрациях в хлоропластах и апопласте. Защитное действие глутатиона связано с окислением его SH-группы, приводящим к димеризации в дисульфид. Антиоксидантные свойства аскорбата основаны на функционировании одноэлектронных циклических переходов между гидро-, семидегидро- и дегидроаскорбатными формами, чему способствует подвижность протонов. Скорость этих превращений зависит от металлов переменной валентности, других пар окислитель-восстановитель и рН среды.
Антиоксидантный эффект аскорбиновой кислоты и глутатиона главным образом реализуется посредством их участия в работе ферментативных антиоксидантов, особенно ферментов аскорбат-глутатионового цикла. Кроме того, они способны непосредственно ингибировать АФК, легко перехватывая O2–, НO2., RO2·, HO·, O2 и др.
Аскорбиновая кислота (витамин С) и восстановленный глутатион (GSH), являясь гидрофильными антиоксидантами, находятся в водной фазе клетки, в хлоропластах, митохондриях и других структурах, а также в межмембранном пространстве клеточных органелл.
Глутатион, являясь полифункциональным соединением, представляет собой основной восстановитель клетки. Как и аскорбиновая кислота, он восстанавливает токофероксильные радикалы, а также участвует в обезвреживании вторичных метаболитов окислительного обмена.
Важной функцией водорастворимых соединений является их роль в восстановлении других низкомолекулярных АО. Так, клеточный GSH участвует в поддержании пула восстановленного аскорбата; аскорбиновая кислота способна восстанавливать мембранно-связанный токоферол, а также используется как высокоспецифичный донор в реакции эпоксидации виалоксантина.
Кроме непосредственно антиоксидантного действия, глутатион и аскорбат выполняют многочисленные функции в регуляции метаболизма растений, в частности, участвуют в поддержании окислительно-восстановительного потенциала.
Антиоксиданты липидной фазы представлены двумя основными классами низкомолекулярных соединений. К первому относят фенольные токоферолы и близкие к ним по строению убихиноны и витамин К. Гидрофобные антиоксиданты предохраняют от разрушения мембраны. Так, локализующийся в мембранах α-токоферол (витамин Е) вызывает обрыв цепей свободнорадикального окисления путем взаимодействия с пероксильными и алкоксильными радикалами. Таким способом токоферол нейтрализует свободные радикалы мембранных липидов, перехватывая и устраняя их. Углеводородная цепь токоферола, встраиваясь в мембрану, образует ассоциации с ненасыщенными жирными кислотами мембранных липидов. Токоферол взаимодействует с О2· и 1О2. Кроме того, токоферолы снижают проницаемость мембран, а также связывают свободные жирные кислоты, избыток которых дестабилизирует мембранную структуру.
Помимо токоферола высокую антиокислительную активность проявляют другие фенольные соединения, в частности полифенолы, служащие донорами водорода. Многие из них способны связывать металлы переменной валентности, которые служат катализаторами свободнорадикального окисления. Из полифенолов наиболее известны как антиоксиданты различные флавоноиды.
К липофильным антиокислителям относятся каротиноиды, которые наиболее эффективны в тушении избыточной энергии триплетных хлорофиллов и синглетного кислорода. Воспринимая энергию возбуждения, они рассеивают ее в виде тепла, предотвращая тем самым возможность образования синглетного кислорода. Кроме того, каротиноиды могут взаимодействовать с органическими радикалами жирных кислот, действуя при этом в качестве не доноров водорода, а «ловушек» радикалов. В итоге цепь окисления обрывается. Антиокислительное действие каротиноидов, видимо, дополняет действие токоферолов, поскольку каротиноиды эффективны в субклеточных фракциях с низким уровнем кислорода, а токоферолы – с более высоким. Возможны и прямые взаимодействия между каротиноидами и токоферолами.
Эти соединения в основном перехватывают перекисные и алкоксильные радикалы, но могут ингибировать также O2–, НO2·, 1O2, ОН·. Взаимодействие фенольных АО с органическими радикалами приводит к образованию феноксильных радикалов, которые в дальнейшем могут образовать хинолидные перекиси. Распад последних приводит к образованию хинонных форм молекул, которые не обладают антиоксидантными свойствами и восстанавливаются с помощью других АО, в частности, аскорбата. Фенольные АО также участвуют в ингибировании или инактивации ферментов, активирующих кислород. Механизмы этих процессов могут быть основаны на конкуренции фенолов с субстратом при связывании с регуляторным центром фермента или инактивации активного центра.
Второй класс липорастворимых АО – ретинол, его предшественники и производные. Антиоксидантная роль каротиноидов проявляется в основном в ингибировании синглетного кислорода, но они могут обезвреживать и пероксирадикалы. Наилучшим эффектом обладают β-каротин и ликопин, способные хелатировать ионы Fe2+, а также тушить триплетное состояние хлорофилла, являющегося одним из источников образования 1O2.
Скорость ПОЛ существенно зависит от структурной организации фосфолипидов мембран: чем плотнее их упаковка, тем меньше скорость окисления. Способностью стабилизировать липиды мембран обладают токоферолы, витамин К, убихиноны, каротиноиды, другие соединения. Встраиваясь боковыми цепями между ненасыщенными жирными кислотами (НЖК) мембран, эти вещества образуют комплексы с их двойными связями.
Системы антиоксидантной защиты. Растения обладают достаточной устойчивостью к окислительным повреждениям, которые возникают лишь при резком изменении физиологического состояния организма. Это обусловлено существованием в растительной клетке эффективных антиоксидантных систем (АОС), способных обеспечить защиту как от кислородных радикалов, так и от синглетного кислорода.
Как восстанавливающий агент, аскорбиновая кислота способна напрямую реагировать с супероксид- и гидроксильным радикалами, однако основная ее функция в процессе ПОЛ состоит в регенерации токофероксильного радикала: ТО· + АКН → ТОН + А· (АКН – аскорбиновая кислота, А· – радикал монодегидроаскорбиновой кислоты). Дальнейшее окисление монодегидроаскорбата приводит к образованию дегидроаскорбата. Окисленные формы аскорбиновой кислоты восстанавливаются в ферментативных реакциях монодегидроаскорбат- и дегидроаскорбатредуктазами.
Глутатион представляет собой основной восстановитель клетки. Окисление сульфогруппы глутатиона (Г-SH) приводит к образованию окисленного глутатиона (T-SS-Г), в котором две молекулы Г-SH связываются, образуя дисульфидный мостик. В ходе окислительного стресса количество Г-SS-Г резко увеличивается и вслед за этим активируется синтез восстановленных форм глутатиона. Глутатион – полифункциональное соединение. Как и аскорбиновая кислота, он восстанавливает токофероксильные радикалы: ТО· + Г-SH → ТОН+Г-S·; 2Г-S· + НАДФН → Г-SS-Г + НАДФ+. Он участвует также в восстановлении Н2O2, RООН, обезвреживании вторичных метаболитов окислительного обмена. Глутатион-зависимые ферменты работают во всех частях клетки. Антиоксидантный эффект селена также опосредован глутатионпероксидазами.
Гидрофобные антиоксиданты предохраняют от разрушения мембраны. Локализующийся в мембранах α-токоферол (витамин Е) вызывает обрыв цепей свободнорадикального окисления путем взаимодействия с пероксильными и алкоксильными радикалами: ТОН + ROO· → ТО· + RООН; ТОН + RО· → ТО· + ROH (ТОН – токоферол, ТО· – токофероксильный радикал). Образовавшийся токофероксильный радикал восстанавливается до токоферола или вступает в реакцию с новыми пероксильными радикалами: ТО· + ROO· → молекулярные продукты. При этом образуется большое число молекулярных продуктов с эпокси-, кето-, гидроперокси- и другими группами. Таким образом токоферол нейтрализует свободные радикалы мембранных липидов, перехватывая и устраняя их. Углеводородная цепь токоферола, встраиваясь в мембрану, образует ассоциации с ненасыщенными жирными кислотами мембранных липидов. Токоферол взаимодействует с О2· и 1О2. Кроме того, токоферолы снижают проницаемость мембран, а также связывают свободные жирные кислоты, избыток которых дестабилизирует мембранную структуру.
Помимо токоферола высокую антиокислительную активность проявляют другие фенольные соединения, в частности полифенолы как доноры водорода. Многие из них способны связывать металлы переменной валентности, которые служат катализаторами свободнорадикального окисления. Из полифенолов наиболее известны как антиоксиданты различные флавоноиды.
Основную роль в снижении уровня АФК играют ферментные АОС: СОД, каталаза, пероксидаза, глутатионтрансфераза, фосфолипидгидропероксидаза и др. СОД катализирует реакцию восстановления O2·: O2· + O2· + 2Н+ → Н2O2 + O2, т. е. препятствует накоплению супероксида кислорода. СОД растений содержит в активных центрах ионы меди, цинка, железа или марганца. Cu-Zn-СОД локализована в основном в цитоплазме, хлоропластах и пероксисомах эукариот, Мn-СОД – в митохондриальных мембранах, а Fe-СОД – в хлоропластах большинства растений. Механизм взаимодействия СОД с супероксидным радикалом точно не выяснен. Предполагается, что сначала одна молекула супероксида взаимодействует с активным центром фермента, при этом металл, входящий в активный центр, восстанавливается, а супероксидный радикал окисляется до молекулярного кислорода:
Cu2+ + O2·→ Cu+ + O2.
Затем при участии второй молекулы супероксидного радикала происходит обратное окисление металла, при этом супероксид восстанавливается до пероксида водорода:
Сu+ + O2· + 2Н+ → Сu2+ + Н2O2.
В детоксикации пероксида водорода 2Н2O2 → 2Н2O + O2 важную роль играют Fe-содержащие ферменты: каталаза и различные пероксидазы. Активность каталазы связана в основном с митохондриями, цитоплазмой и пероксисомами. В отличие от пероксидаз, которые функционируют только при относительно низком уровне пероксида, каталаза способна эффективно работать при высокой его концентрации.
Пероксидазы – обширная группа ферментов, катализирующих реакции окисления органического и неорганического субстрата с использованием пероксида водорода или органических пероксидов в качестве акцепторов электронов:
2XH+ Н2O2 → 2Х + 2Н2O;
2XH + RООН → 2X + H2O + ROН,
где XH – восстановленный субстрат, X– окисленный субстрат. Функции пероксидаз разнообразны. Они участвуют в синтезе лигнина, связаны с процессами роста и растяжения, участвуют в катаболизме ауксина и т. д. К важным функциям пероксидаз относится участие их в защите растения-хозяина от патогена, катализ окисления фенолов до хинонов, а также образование супероксида-аниона и пероксида водорода (оксидазная функция). Антиоксидантную защиту, связанную с восстановлением пероксида водорода, осуществляют главным образом глутатионпероксидаза и аскорбатпероксидаза. Кроме того, в вакуолях растений, а также у организмов с низкой активностью аскорбатпероксидазы, детоксикация пероксидов эффективно обеспечивается гваяколпероксидазой.
Глутатионпероксидаза, локализованная в цитоплазме, плазмалемме и в матриксе митохондрий, разрушает пероксид водорода и органические гидропероксиды свободных жирных кислот, нуклеотидов, нуклеиновых кислот, белков: Н2O2 + 2ГSH → 2Н2O + ГSSГ; ROОН + 2ГSН → ROН + Н2O + ГSSГ. Среди глутатионпероксидаз выделяют связанную с мембранами селен-зависимую форму, катализирующую в основном редукцию гидропероксидов фосфолипидов. Эта фосфолипидгидроксипероксид-глутатионпероксидаза восстанавливает ROOН жирных кислот в составе фосфолипидов. В строме хлоропластов образуется много пероксида водорода, и в его элиминировании участвует несколько сопряженных ферментативных систем.
Пероксид водорода разрушается аскорбатпероксидазой, две формы которой локализованы в хлоропластах, а две другие – в мембранах глиоксисом и в цитоплазме. Окисленный аскорбат восстанавливается аскорбатредуктазой, в реакции участвует восстановленный глутатион. В свою очередь, глутатион регенерируется глутатионредуктазой, в результате образуется ГSH из ГSSГ путем НАДФН-зависимого восстановления: ГSSГ + НАДФН + Н+ → 2ГSH + НАДФ+. Эта реакция уменьшает или даже предупреждает прогрессирование ПОЛ и окислительный распад нуклеиновых кислот и белков. Глутатионтрансфераза восстанавливает только ROОН, причем один из ее изоферментов находится в хроматине и восстанавливает ROОН ДНК в ядре.
Гваяколпероксидазы, или «классические пероксидазы», катализируют окисление большого набора ароматических соединений с использованием пероксида водорода или органических пероксидов в качестве акцепторов электронов. Помимо участия гваяколпероксидаз в процессах лигнификации, распада ауксина и др. им отводят важную роль и в защите клетки от окислительного стресса. Например, в вакуолях или клеточной стенке, не имеющих аскорбатпероксидаз, но содержащих в достаточно высоких концентрациях фенолы и аскорбиновую кислоту, может функционировать комплекс гваяколпероксидаза-фенолы-аскорбиновая кислота. При этом фенолы окисляются гваяколпероксидазой, а затем восстанавливаются аскорбиновой кислотой, в результате происходит детоксикация продуктов окисления фенолов. Таким образом, в клеточных стенках и вакуолях происходит эффективное обезвреживание пероксида водорода, что предотвращает возможность проникновения его в другие субклеточные структуры.
Вторичные метаболиты окислительных модификаций также обезвреживаются различными ферментами: глутатионтрансфераза катализирует конъюгацию с ГSH ряда окисленных веществ, формальдегиддегидрогеназа и глиоксилаза, использующие ГSH в качестве кофермента, окисляют свои субстраты до органических кислот, альдегиддегидрогеназа окисляет МДА, хинонредуктаза восстанавливает хиноны до дегидрохинонов, что предупреждает образование продуктов одноэлектронного восстановления семихинонов. Таким образом, ферментативная АОС обеспечивает разрушение не только АФК, но и их производных окисленных соединений.
При дефиците низкомолекулярных антиоксидантов – глутатиона, витаминов Е и С – накапливаются АФК и нарастает содержание окисленных веществ, т. е. окислительный стресс возникает не только при избытке АФК и оксидативных модификаций макромолекул, но и при недостатке АОС (не только абсолютного, но и относительного). Следовательно, окислительный стресс – это сдвиг к преобладанию прооксидантов над антиоксидантами. Соотношение прооксидантов и антиоксидантов лежит в основе внутриклеточной редокс-регуляции. Эта концепция подразумевает признание регуляторных функций АФК и АОС. Итак, в регуляции биологических процессов, по всей вероятности, следует выделить новую форму контроля – редокс-регуляцию, или контроль соотношения прооксидантов и антиоксидантов, что, по сути, и определяет, будет прогрессировать оксидативный стресс или удастся его блокировать.
Образование АФК и последующие оксидативные изменения органических веществ представляют собой неспецифические реакции. У растений, устойчивых к различным воздействиям, процессы генерации АФК обычно заторможены, что во многом определяется активным функционированием у них систем детоксикации. Возможно, именно с этим связана в значительной мере и большая стойкость мембран устойчивых растений к стрессорам. Устойчивые растения обладают более высоким содержанием аскорбата, α-токоферола, активностью антиоксидантных ферментов. В неблагоприятных условиях растения способны к индукции активности АОС, в результате они приобретают устойчивость не только к действующему агенту (например, гербициду), но и к другим воздействиям, вызывающим образование АФК. Такие факты послужили стимулом для создания с помощью методов генной инженерии форм растений, обладающих повышенной устойчивостью к гербицидам, инфекционным заболеваниям, загрязнителям атмосферы и др. Известны сообщения о создании таких организмов, которые имеют очень высокую активность СОД, аскорбатпероксидазы, глутатионредуктазы. Трансгенные растения, у которых была экспрессирована СОД, оказывались более толерантными к водному дефициту, низкой температуре. Инфильтрация в ткани СОД подавляла как генерацию АФК, так и гибель пораженных патогеном клеток. Вместе с тем у растений, акклиматизированных к пониженной температуре, повышалась устойчивость к гербицидам, образующим АФК. Эти данные подтверждают рассмотренные выше представления о сопряженной устойчивости растений. Они свидетельствуют также в пользу гипотезы о том, что устойчивость растений к окислительному стрессу выступает важным фактором в приспособлении организмов к различным экстремальным воздействиям.
Контрольные вопросы и задания
- Охарактеризуйте активные формы кислорода.
- Каково биологическое значение АФК?
- Какие стрессовые воздействия могут привести к избыточным концентрациям АФК?
- Охарактеризуйте компартментацию АФК в клетке.
- Как повреждаются биополимеры активными формами кислорода?
- Охарактеризуйте методы оценки локализации окислительного стресса в растении.
- В чем состоят механизмы детоксикации АФК?
- Дайте классификацию антиоксидантов в зависимости от их молекулярной массы и растворимости.
- Какую роль играют антиоксиданты?
- Какую роль играют каротиноиды и флавоноиды в реакции на окислительный стресс?
- Каковы условия образования АФК?
- Какую роль выполняет озон в генерировании АФК?
- В чем состоит антиоксидантный эффект аскорбиновой кислоты и глутатиона?
- Каковы основные пути антиоксидантной защиты растений?
Глава 3. ДЕЙСТВИЕ ДЕФИЦИТА ВОДЫ И ЗАСУХОУСТОЙЧИВОСТЬ
Более 70% пресной воды, используемой человечеством, приходится на потребности сельского хозяйства, и эта цифра будет только возрастать. В США, например, более 40% страховых выплат в сельском хозяйстве приходится именно на потери урожая при засухе (Bacon, 2004). Одним из путей решения проблемы является выведение засухоустойчивых сортов с высокой эффективностью транспирации.
3.1. Эффективность использования воды растениями
Для оценки эффективности использования воды (ЭИВ) используют следующие показатели:
1) ассимиляционно-транспирационное отношение (АТО), рассчитываемое как соотношение текущих значений интенсивности фотосинтеза и транспирации листьев;
2) продуктивность транспирации (ПТ), определяемая как соотношение сухой биомассы (вегетативной или хозяйственно-полезной), сформированной на единицу суммарных потерь воды за счет транспирации и испарения с поверхности почвы (эвапотранспирация). Данный показатель, в отличие от АТО, рассчитывается за весь период вегетации и более корректно характеризует ЭИВ;
3) метод дискриминации углерода (Δ13С), предложенный группой австралийских ученых для C3 растений (Farguhar et al., 1982), характеризует АТО на уровне отдельного листа за весь вегетационный период, косвенно отражая продуктивность транспирации. Метод основан на дискриминации поглощения более тяжелого (13С) изотопа углерода по сравнению с 12С при фотосинтезе, в результате чего соотношение 13С/ 12С в сухой массе оказывается ниже, чем в воздухе. Величина дискриминации Δ13С зависит от генотипа, и она меньше, когда АТО выше (рис. 3.1).
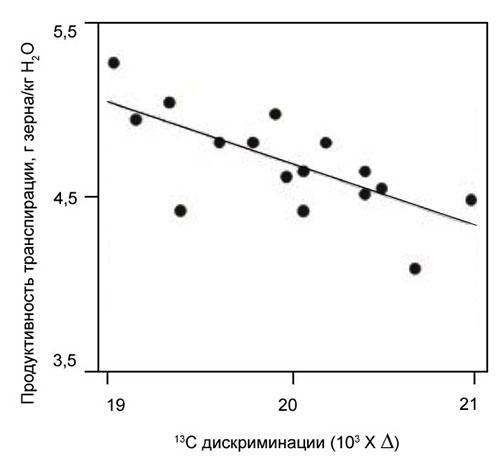
Рис. 3.1. Связь между ∆13С и продуктивностью транспирации у разных генотипов пшеницы при оптимальной влагообеспеченности (Condon et. al., 1990)
Использование Δ13С в целях селекции, в частности пшеницы, осложнено зависимостью дискриминации 13С от условий окружающей среды, а также необходимостью наличия масс-спектрометра и высокой стоимостью анализов (5–15$ за образец). Кроме того, искомый метод не позволяет вычленить дыхательные потери СО2 и испарение воды из почвы, а также выяснить, что является причиной вариации АТО – изменение ИФ или устьичной проводимости. Параллельные измерения дискриминации изотопа кислорода (18О) могут помочь в решении ряда указанных проблем, так как содержание 18О в органическом материале листа непосредственно связано со скоростью испарения. Это объясняется тем, что вода в листе обогащается 18О в результате того, что легкий изотоп 16О предпочтительно испаряется в ходе процесса транспирации.
Таким образом в основе всех отмеченных выше показателей ЭИВ находится сопряженность процессов газо- и водообмена на разных уровнях организации растения.
3.1.1. Регуляция листом газо- и водообмена
Устьичный контроль и проводимость ксилемы. Механизм устьичной реакции на засуху достаточно сложен, поскольку в каждый конкретный момент устьица могут реагировать на комплекс факторов, включающий уровень освещенности, концентрацию СО2, температуру и водный статус листа.
При этом накапливается все больше данных, указывающих на то, что закрывание устьиц опосредовано химическими сигналами, исходящими в направлении побегов от теряющих воду корней. Кроме уже известной роли АБК в закрывании устьиц, другие гормоны, видимо, тоже участвуют в этом процессе или по отдельности, или во взаимодействии с АБК. Именно последняя была идентифицирована в качестве одного из химических сигналов, участвующих в регуляции функционирования устьиц, достаточно сложного процесса, включающего как дальний транспорт, так и модуляцию концентрации АБК в замыкающих клетках устьиц. К факторам указанной модуляции относятся рН ксилемного сока и листовой ткани, которые могут возрасти в условиях, вызывающих ускоренную транспирацию (высокий дефицит давления водяных паров в воздухе, высокие освещенность и температура листа). Закрывание устьиц после полудня при оптимальном влагообеспечении тоже можно частично объяснить повышением рН. Вместе с тем о сложности реакции растений на АБК свидетельствует тот факт, что формы, отличающиеся высоким содержанием АБК, далеко не всегда засухоустойчивы.
Важная роль, которую устьица играют в компромиссе между «жаждой» и «голодом» (соответственно потерей воды при транспирации и поглощением СО2 при фотосинтезе), обусловлена их исключительной чувствительностью как к условиям среды, так и к внутренним физиологическим факторам. Например, полуденная депрессия устьичной проводимости в полевых условиях является важной регуляторной реакцией для контроля за потерей воды (даже в условиях нелимитированного водообеспечения) и оптимизации АТО. Величина депрессии зависит от влажности и температуры воздуха, водного статуса растения, а также засухоустойчивости сорта. Есть данные, что причиной полуденного закрывания устьиц является не изменение концентрации АБК или скорости тока воды в ксилеме, а увеличение рН ксилемного сока в течение дня (рис. 3.2). В качестве другой причины полуденной депрессии газообмена листа называется реакция устьиц на избыточную солнечную радиацию.
Однако независимо от причин закрывания устьиц, большинство видов культурных растений демонстрируют увеличение эффективности использования воды (ЭИВ) при умеренном водном дефиците. Это обусловлено нелинейной связью между ИФ и гидравлической проводимостью устьиц (gs), т. е. интенсивность транспирации (ИТ) ингибируется раньше и сильнее, чем ИФ. Это заметнее проявляется на С4-, чем на С3-видах, поскольку у С4-видов поглощение СО2 менее чувствительно к начальному снижению устьичной проводимости, чем у С3-видов.
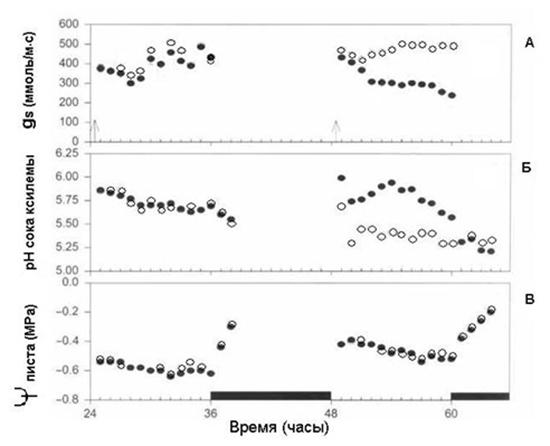
Рис. 3.2. Зависимость устьичной проводимости (А), рН ксилемного сока (Б) и водного потенциала листа (В) растений томата, выращиваемых по технологии расщепленных корней, от частоты полива. Закрашенные круги – полив одной половины корней, незакрашенные круги – полив всех корней, Стрелки указывают время полива, а выделенная часть на оси времени – ночной период. Влияние дефицита влаги на устьичную проводимость заметно на третьи сутки, и оно совпадает с подщелачиванием ксилемного сока (Bacon, 2004)
Устьичная аппертура играет заметную роль в терморегуляции листа. За длительный период селекции на высокую урожайность при повышенных температурах gs повышалась параллельно с урожайностью хлопчатника и пшеницы при орошении, что позволяет использовать устьичную проводимость в качестве важного селекционного параметра. Поскольку молекулы СО2 и Н2О используют один и тот же путь диффузии, но диффузный градиент, управляющий транспирацией, заметно больше (примерно в 50 раз), чем для поглощения СО2, увеличение gs неизбежно приводит к более существенному увеличению ИТ. При этом устьица открыты в той мере, какая необходима, чтобы обеспечить поглощение СО2 при фотосинтезе, одновременно избегая обезвоживания тканей в процессе транспирации. При этом отношение ИФ к устьичной проводимости остается достаточно постоянным в широком диапазоне действия факторов (табл. 3.1).
Таблица 3.1
Интенсивность фотосинтеза (ИФ, мкмоль СО2/м2 · сек), устьичная проводимость (gs, ммоль/м2 · сек) и их соотношение (ммоль СО2 /моль Н2О) утром и в полдень при различной температуре (Bacon, 2004)
| Показатели | Утро | Полдень | ||
| 22,5 °С | 32,5 °С | 22,5 °С | 32,5 °С | |
| ИФ | 11,6 | 4,8 | 7,8 | 3,2 |
| gs | 134,6 | 57,7 | 84,6 | 42,3 |
| ИФ/ gs | 86,2 | 83,2 | 92,2 | 75,7 |
Этот вывод положен в основу концепции, согласно которой эволюция растений сопровождалась оптимизацией водопользования. При этом парциальное давление СО2 внутри клетки и АТО остаются постоянными, за исключением резкого изменения факторов среды, причем АТО в каждом конкретном случае достигает максимальной для данных условий выращивания величины.
При длительном водном стрессе эффективность использования воды, характеризуемая в данном случае АТО, может коррелировать с относительной скоростью роста (ОСР) как положительно, так и отрицательно. Первое, как правило, характерно для С4-видов, избегающих действия засухи за счет быстрого роста в начале вегетации. Отрицательная корреляция между АТО и ОСР установлена для засухоустойчивых видов, причем повышение АТО при засухе сочетается с перераспределением большей доли ассимилятов в нефотосинтезирующие структуры, например, корни. Подобная стратегия характерна для медленнорастущих видов с длинным периодом вегетации, и повышенное АТО в данном случае обусловлено экономным использованием воды и необходимостью выживания.
3.1.2. Функционирование корневой системы
Удельная длина и распределение корней в почве. Насыщенность почвы корнями характеризуется удельной длиной корней (УДК), равной длине корней в единице объема почвы (см/см3). Однодольные культуры, в отличие от двудольных, обнаруживают более высокие значения УДК. Так, в профиле почвы глубиной 1,3 м УДК для кукурузы составила в среднем 0,22, а для сои – 0,08 см/см3. Гораздо более высокие и вариабельные значения получены в другом опыте, где сравнивали кукурузу (3,7–6,2 см/см3) и хлопчатник (1,1–1,8 см/см3). Таким образом, даже в пределах одного вида значения УДК заметно различаются (см.
...


















